Базовый слой
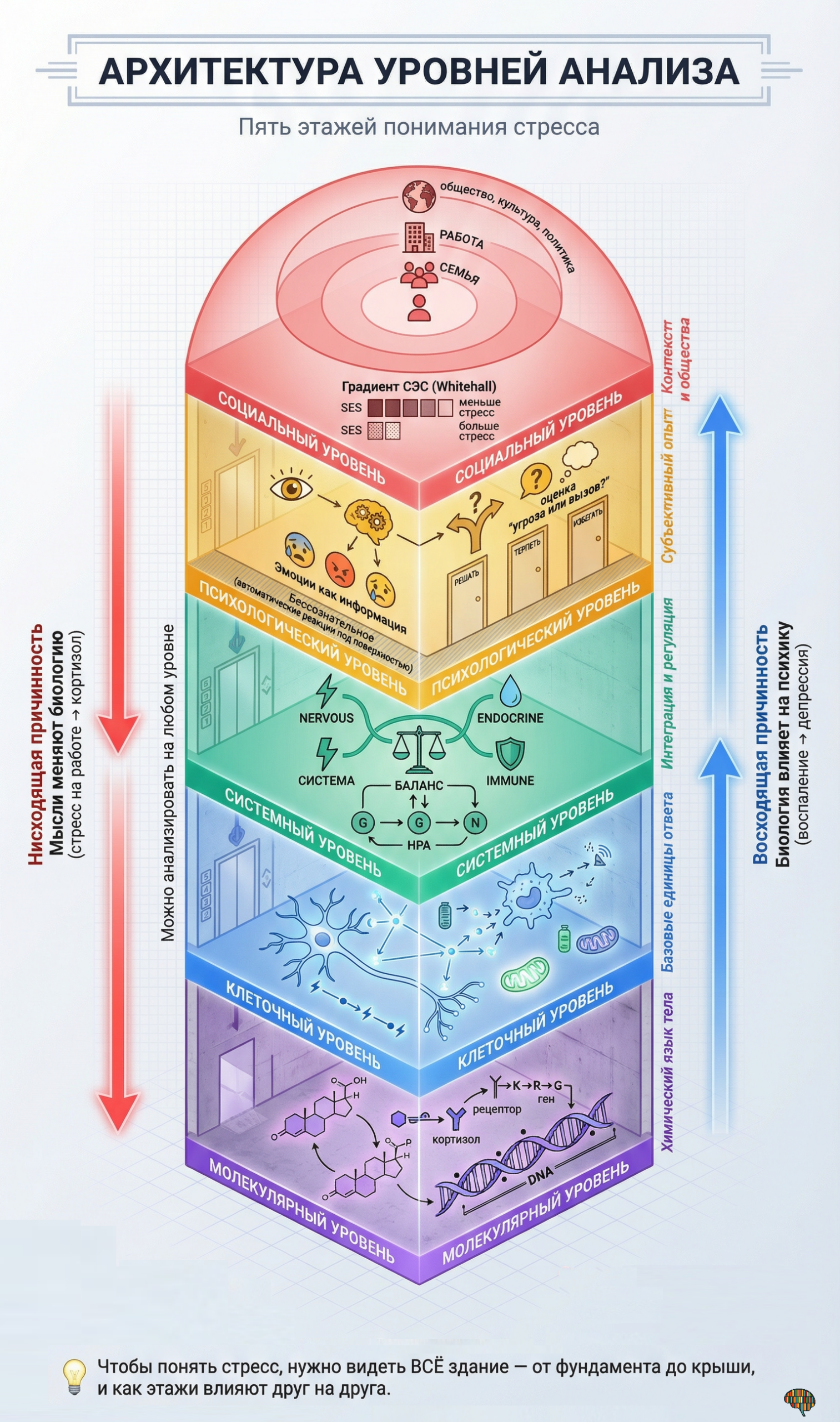
1. Молекулярный уровень — невидимый язык стресса
1.1 Химические посредники как язык организма
Человеческий организм представляет собой сложнейшую систему, в которой триллионы клеток должны действовать согласованно, чтобы обеспечить выживание и адаптацию к постоянно меняющимся условиям среды. Эта координация невозможна без непрерывного обмена информацией между клетками, тканями и органами, и именно молекулы выступают в роли универсального языка, на котором различные части организма общаются друг с другом. Когда мы говорим о стрессе на молекулярном уровне, мы в первую очередь обращаемся к этой системе химической коммуникации, которая позволяет организму мгновенно мобилизовать ресурсы в ответ на угрозу и координировать сложнейшие физиологические изменения во множестве систем одновременно. Понимание этого молекулярного диалога составляет фундамент современной науки о стрессе, поскольку все более высокие уровни анализа — клеточный, системный, психологический и социальный — в конечном счёте реализуются через изменения в концентрации и активности специфических молекул. Без этого понимания любые рассуждения о стрессе остаются поверхностными и лишёнными механистической основы.
Концепция молекулярной коммуникации в организме развивалась постепенно на протяжении всего двадцатого века, начиная с открытия первых гормонов и заканчивая современным пониманием сложнейших сигнальных сетей. Термин «гормон» был введён в начале прошлого столетия для обозначения веществ, которые вырабатываются в одной части тела и оказывают влияние на другие, отдалённые органы, путешествуя по кровеносной системе. Однако со временем стало ясно, что химическая коммуникация в организме значительно разнообразнее и включает не только классические гормоны, но и нейромедиаторы, действующие в синаптических щелях между нейронами, цитокины, координирующие иммунный ответ, локальные медиаторы, влияющие на соседние клетки, и даже внутриклеточные сигнальные молекулы, передающие информацию от мембраны к ядру клетки. Все эти разнообразные классы химических посредников участвуют в стресс-ответе, создавая многослойную систему коммуникации, которая обеспечивает как скорость реакции, так и её точную настройку под конкретные обстоятельства.
Метафора языка применительно к молекулярной коммуникации оказывается на удивление точной и продуктивной для понимания принципов работы этой системы. Подобно тому как человеческий язык состоит из ограниченного набора слов, которые комбинируются для создания бесконечного разнообразия сообщений, организм использует относительно ограниченный набор сигнальных молекул, но их комбинации, концентрации и временные паттерны позволяют передавать чрезвычайно разнообразную информацию. Одна и та же молекула может нести разные «сообщения» в зависимости от контекста: её концентрации, продолжительности воздействия, типа клетки-получателя и присутствия других сигнальных молекул. Например, кортизол в низких концентрациях и при кратковременном воздействии передаёт сообщение о необходимости мобилизации энергетических ресурсов, тогда как хронически повышенный уровень того же самого гормона несёт совершенно иное послание, связанное с подавлением иммунитета и перестройкой метаболизма в режим долгосрочного выживания. Эта контекстуальная зависимость значения молекулярных сигналов представляет собой одну из ключевых особенностей биологической коммуникации.
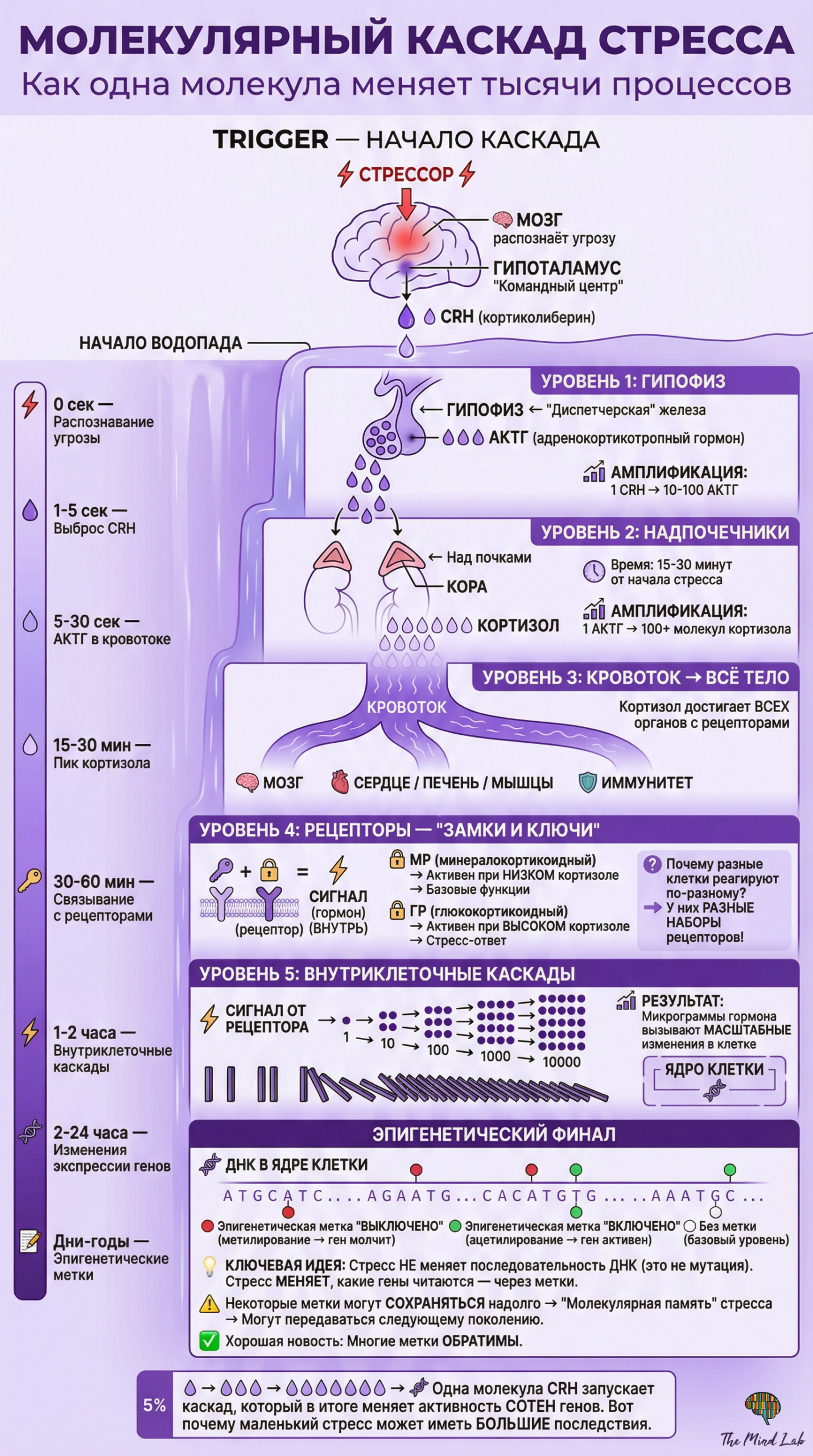
Процесс передачи молекулярного сообщения при стрессе разворачивается по определённому алгоритму, который включает несколько обязательных этапов. Всё начинается с восприятия стрессора — будь то физическая угроза, социальный конфликт или внутреннее нарушение гомеостаза — специализированными сенсорными системами организма. Эта информация поступает в мозг, где происходит её интеграция и оценка, после чего командные центры, прежде всего гипоталамус, инициируют выброс первичных сигнальных молекул. Эти молекулы путешествуют к своим мишеням — либо по нервным волокнам в виде электрических импульсов, преобразуемых в химические сигналы в синапсах, либо через кровоток в случае гормонов. Достигнув клеток-мишеней, сигнальные молекулы взаимодействуют со специфическими рецепторами, что запускает внутриклеточные каскады, ведущие к изменению активности клетки. Важно понимать, что этот процесс не является односторонним: клетки-мишени посылают обратные сигналы, информируя командные центры о своём состоянии, что позволяет организму точно регулировать интенсивность стресс-ответа и вовремя его прекращать.
Когда человек сталкивается с угрозой — например, внезапно видит приближающийся автомобиль — молекулярный ответ разворачивается с поразительной скоростью и точностью. В течение долей секунды зрительная информация достигает миндалевидного тела, которое оценивает её как потенциально опасную ещё до того, как возникает сознательное понимание ситуации. Миндалина немедленно активирует гипоталамус, который посылает сигналы по двум параллельным путям. Первый, быстрый путь задействует симпатическую нервную систему и приводит к выбросу адреналина из мозгового вещества надпочечников буквально в течение секунд. Второй, более медленный путь активирует гипоталамо-гипофизарно-надпочечниковую ось, что приводит к выбросу кортизола в течение нескольких минут. Одновременно в мозге изменяется баланс нейромедиаторов: повышается активность норадреналиновой системы, усиливающей бдительность, и снижается активность серотониновой системы, что может способствовать более импульсивному поведению. Все эти молекулярные изменения скоординированы во времени и пространстве таким образом, чтобы обеспечить максимально эффективный ответ на угрозу.
Особенностью молекулярной коммуникации при стрессе является её каскадный характер, когда небольшое количество первичных сигнальных молекул способно вызывать масштабные изменения в работе всего организма. Это достигается благодаря механизмам амплификации сигнала, при которых одна молекула гормона, связавшись с рецептором, запускает синтез десятков или сотен вторичных посредников внутри клетки, каждый из которых, в свою очередь, активирует множество эффекторных молекул. Такая организация позволяет организму реагировать на стресс с минимальными энергетическими затратами на производство сигнальных молекул, одновременно обеспечивая мощный и быстрый ответ. Однако каскадная организация имеет и обратную сторону: она создаёт множество точек, в которых регуляция может нарушиться, что при хроническом стрессе ведёт к патологическим изменениям, поскольку небольшие отклонения на ранних этапах каскада могут многократно усиливаться на последующих этапах.
Клетки организма не являются пассивными получателями молекулярных сообщений — они активно «слушают» и «отвечают» на химические сигналы, формируя двунаправленную коммуникацию. Каждая клетка экспрессирует определённый набор рецепторов, который определяет, какие сигналы она способна воспринимать, и этот набор может динамически изменяться в зависимости от состояния клетки и окружающей среды. При повторяющемся воздействии сигнальных молекул клетка может уменьшать количество рецепторов на своей поверхности — феномен, известный как даунрегуляция, — что делает её менее чувствительной к данному сигналу. Этот механизм лежит в основе привыкания и толерантности, но при хроническом стрессе он может вести к нарушению нормальной регуляции. Помимо этого, клетки сами выделяют сигнальные молекулы, которые влияют на их соседей и могут достигать отдалённых органов, создавая сложные петли обратной связи. Понимание этих механизмов клеточного ответа критически важно для объяснения того, почему одинаковый стрессор может вызывать различные реакции у разных людей или у одного человека в различных состояниях.
Современные методы исследования позволяют изучать молекулярную коммуникацию при стрессе с беспрецедентной детальностью, открывая всё новые слои сложности этой системы. Технологии масс-спектрометрии позволяют одновременно измерять концентрации сотен метаболитов в образце крови или ткани, создавая «молекулярный портрет» стрессового состояния. Методы транскриптомики показывают, какие гены активируются или подавляются в ответ на стресс в различных типах клеток. Протеомика выявляет изменения в составе и модификациях белков. Интеграция этих данных в рамках системной биологии позволяет строить всё более полные карты молекулярных событий, разворачивающихся при стрессе. Однако даже с этими мощными инструментами мы лишь начинаем понимать всю сложность молекулярного языка организма, и каждое новое открытие ставит новые вопросы о механизмах координации и регуляции этой удивительной системы коммуникации.
Значение понимания молекулярного уровня стресса выходит далеко за пределы чисто академического интереса и имеет прямые практические следствия для медицины, психологии и общественного здравоохранения. Именно на молекулярном уровне действуют лекарственные препараты, применяемые для лечения стресс-связанных расстройств: антидепрессанты изменяют баланс нейромедиаторов, анксиолитики воздействуют на рецепторы тормозных медиаторов, противовоспалительные средства блокируют сигнальные пути цитокинов. Понимание молекулярных механизмов также объясняет, почему психологические интервенции — когнитивная терапия, медитация, социальная поддержка — оказывают реальное влияние на физическое здоровье: все эти воздействия в конечном счёте реализуются через изменения в молекулярной среде организма. Более того, молекулярные биомаркеры стресса открывают возможности для ранней диагностики и персонализированного подбора терапии. Таким образом, молекулярный уровень анализа представляет собой не просто один из многих возможных взглядов на стресс, но фундаментальный уровень, на котором все остальные аспекты стресса находят своё материальное воплощение.
1.2 Гормоны стресса — первое знакомство с главными героями
Среди всего многообразия молекул, участвующих в стресс-ответе, особое место занимают гормоны — химические посредники, которые синтезируются специализированными железами и путешествуют по кровеносной системе к своим мишеням, оказывая системное влияние на работу множества органов. История изучения гормонов стресса неразрывно связана с именами пионеров эндокринологии и физиологии стресса, начиная с Уолтера Кэннона, описавшего реакцию «бей или беги», и Ганса Селье, сформулировавшего концепцию общего адаптационного синдрома. За прошедшее столетие наше понимание гормональной регуляции стресса многократно углубилось, однако три молекулы по-прежнему остаются центральными действующими лицами этой драмы: кортизол, адреналин и норадреналин. Каждый из этих гормонов обладает уникальным профилем действия, временной динамикой и набором мишеней, и именно их согласованная работа обеспечивает адаптивный ответ организма на стресс. Знакомство с этими «главными героями» молекулярной драмы стресса необходимо для понимания всех последующих уровней анализа.
Кортизол по праву считается главным гормоном стресса и занимает центральное место в регуляции долгосрочных адаптивных процессов. Этот стероидный гормон синтезируется в корковом веществе надпочечников под контролем гипоталамо-гипофизарно-надпочечниковой оси и относится к классу глюкокортикоидов, названных так из-за их выраженного влияния на метаболизм глюкозы. Кортизол обладает поразительно широким спектром действия, что объясняется присутствием его рецепторов практически во всех типах клеток организма. Его основная функция при стрессе может быть охарактеризована метафорой «мобилизатора ресурсов»: кортизол перераспределяет энергетические субстраты таким образом, чтобы обеспечить выживание организма в условиях угрозы. Он стимулирует глюконеогенез в печени, повышая уровень глюкозы в крови, способствует расщеплению жиров и белков для обеспечения энергетических субстратов, подавляет несущественные для немедленного выживания функции, такие как репродукция и рост, и оказывает сложное модулирующее влияние на иммунную систему.
Временная динамика секреции кортизола существенно отличает его от быстродействующих гормонов стресса и определяет его роль в регуляции пролонгированных адаптивных процессов. После воздействия стрессора концентрация кортизола в крови начинает повышаться лишь через несколько минут и достигает пика примерно через двадцать-тридцать минут, что связано с многоступенчатым характером активации гипоталамо-гипофизарно-надпочечниковой оси. Важно понимать, что кортизол существует в организме не только как инструмент стресс-ответа, но и как регулятор базовых метаболических процессов с выраженным суточным ритмом: его концентрация максимальна в утренние часы, обеспечивая энергетическую мобилизацию для начала активного дня, и минимальна в ночное время, когда преобладают восстановительные процессы. Этот циркадный ритм кортизола имеет критическое значение для здоровья, и его нарушение при хроническом стрессе связано с широким спектром патологических последствий, от метаболических нарушений до депрессии. Понимание двойственной природы кортизола — как гормона стресса и как регулятора суточных ритмов — необходимо для адекватной интерпретации его уровней в клинических и исследовательских контекстах.
Влияние кортизола на мозг заслуживает особого внимания, поскольку именно через эти эффекты реализуется связь между биологическим и психологическим уровнями стресса. Рецепторы кортизола присутствуют в высокой концентрации в гиппокампе, миндалине и префронтальной коре — структурах, критически важных для памяти, эмоциональной обработки и исполнительного контроля. При остром стрессе умеренное повышение кортизола усиливает консолидацию эмоционально значимых воспоминаний, что имеет очевидный адаптивный смысл: опасные ситуации должны хорошо запоминаться для предотвращения их повторения. Однако хронически повышенный уровень кортизола оказывает нейротоксическое действие, особенно на гиппокамп, где он подавляет нейрогенез и может приводить к атрофии дендритов нейронов. Эти структурные изменения лежат в основе когнитивных нарушений, наблюдаемых при хроническом стрессе и депрессии, включая трудности с концентрацией внимания и нарушения памяти. Таким образом, кортизол представляет собой классический пример молекулы с дуальным действием, полезным в острых ситуациях и потенциально разрушительным при хронической активации.
Адреналин, также известный в научной литературе под термином «эпинефрин», представляет собой принципиально иной тип гормона стресса, обеспечивающий молниеносную мобилизацию организма для немедленного действия. Этот катехоламин синтезируется в мозговом веществе надпочечников и выбрасывается в кровь в течение секунд после восприятия угрозы, что обеспечивается прямой нервной активацией надпочечников через симпатическую нервную систему. Метафора «аварийной кнопки» точно передаёт суть физиологической роли адреналина: он обеспечивает немедленную готовность организма к интенсивной физической активности. Под влиянием адреналина учащается сердцебиение и усиливается сила сердечных сокращений, расширяются бронхи, облегчая дыхание, расширяются зрачки, улучшая периферическое зрение, повышается артериальное давление, обеспечивая приток крови к мышцам, мобилизуется глюкоза из печёночных депо, снабжая мышцы топливом. Все эти изменения происходят практически мгновенно и хорошо знакомы каждому, кто когда-либо испытывал сильный испуг: колотящееся сердце, учащённое дыхание, ощущение прилива энергии.
Быстрота действия адреналина объясняется особенностями его молекулярной биологии и отличает его от медленнодействующего кортизола. В то время как кортизол должен проникнуть внутрь клетки, связаться с ядерными рецепторами и изменить экспрессию генов — процесс, требующий десятков минут, — адреналин действует через рецепторы на поверхности клетки, активируя быстрые внутриклеточные сигнальные каскады. Адренорецепторы делятся на альфа- и бета-подтипы, каждый из которых запускает различные внутриклеточные каскады и преобладает в определённых тканях, что объясняет разнообразие эффектов адреналина в различных органах. В сердце преобладают бета-адренорецепторы, стимуляция которых ускоряет ритм и усиливает сокращения. В сосудах кожи и пищеварительного тракта преобладают альфа-рецепторы, вызывающие сужение сосудов и перенаправляющие кровоток к мышцам. Эта тканевая специфичность действия адреналина представляет собой элегантный механизм координации множества физиологических изменений посредством одной сигнальной молекулы.
Продолжительность действия адреналина принципиально отличается от кортизола и ограничивается минутами, что соответствует его роли в обеспечении немедленных реакций. Адреналин быстро инактивируется ферментами и удаляется из кровотока, что позволяет организму быстро вернуться к исходному состоянию после устранения угрозы. Однако при повторяющемся или хроническом стрессе даже эти кратковременные подъёмы адреналина могут кумулятивно воздействовать на сердечно-сосудистую систему, повышая риск гипертонии, аритмий и других патологий. Более того, субъективное переживание действия адреналина — ощущение возбуждения, тревоги, повышенной бдительности — может интерпретироваться как симптом тревоги или паники, создавая порочный круг, в котором физиологическое возбуждение усиливает психологическую тревогу, которая, в свою очередь, стимулирует дальнейший выброс адреналина. Понимание этой связи между молекулярными событиями и субъективным переживанием имеет важное клиническое значение для терапии тревожных расстройств.
Норадреналин, третий ключевой гормон стресса, занимает уникальное положение, функционируя одновременно как гормон и как нейромедиатор в центральной нервной системе. В качестве гормона норадреналин выбрасывается надпочечниками вместе с адреналином и оказывает сходные периферические эффекты, хотя с несколько иным профилем рецепторной активации. Однако его роль как нейромедиатора в мозге, пожалуй, ещё более значима для понимания стресса. Норадренергические нейроны сконцентрированы в небольшом ядре ствола мозга, называемом голубым пятном, откуда их аксоны распространяются практически во все отделы мозга. При стрессе активность голубого пятна резко возрастает, и выброс норадреналина модулирует работу коры, гиппокампа, миндалины и других структур. Метафора «прожектора внимания» хорошо передаёт основную функцию норадреналина: он повышает бдительность, обостряет внимание к потенциально значимым стимулам и усиливает обработку сенсорной информации.
Влияние норадреналина на когнитивные функции подчиняется закономерности, известной как кривая Йеркса-Додсона, которая описывает взаимосвязь между уровнем возбуждения и эффективностью деятельности в форме перевёрнутой буквы U. При низком уровне норадреналина внимание рассеяно, мотивация снижена, когнитивные функции не оптимальны. Умеренное повышение норадреналина при остром стрессе улучшает концентрацию, ускоряет обработку информации, повышает готовность к действию. Однако при очень высоких уровнях, характерных для сильного стресса или паники, когнитивные функции резко ухудшаются: внимание становится туннельным, способность к гибкому мышлению падает, преобладают автоматические, стереотипные реакции. Это объясняет, почему умеренный стресс может повышать продуктивность, тогда как сильный стресс парализует мышление. Понимание этой закономерности имеет практическое значение: оптимальное функционирование требует поддержания уровня активации в срединном диапазоне, не слишком низком и не слишком высоком.
Взаимодействие трёх основных гормонов стресса создаёт координированную систему, в которой каждый компонент выполняет специфическую роль, определяемую его временной динамикой и спектром мишеней. Адреналин обеспечивает немедленную мобилизацию в первые секунды и минуты после восприятия угрозы, подготавливая организм к физическому действию. Норадреналин параллельно модулирует работу мозга, обостряя внимание и готовя когнитивные системы к быстрому принятию решений. Кортизол, действуя с задержкой в минуты и часы, перенаправляет метаболизм, обеспечивает устойчивую энергетическую поддержку и модулирует воспалительные процессы. Если угроза быстро устраняется, адреналин и норадреналин быстро инактивируются, кортизол постепенно снижается благодаря механизмам обратной связи, и организм возвращается к исходному состоянию. Проблемы возникают, когда стрессор персистирует или когда механизмы завершения стресс-ответа нарушены — тогда хроническое воздействие этих гормонов начинает оказывать повреждающее действие на ткани и системы организма, что составляет биологическую основу стресс-связанных заболеваний.
Необходимо подчеркнуть, что адреналин, норадреналин и кортизол, при всей их центральной роли, представляют лишь верхушку айсберга гормональной регуляции стресса. Десятки других гормонов и гормоноподобных веществ участвуют в стресс-ответе, включая кортиколиберин и адренокортикотропный гормон, регулирующие секрецию кортизола, окситоцин и вазопрессин, модулирующие социальное поведение и водно-солевой баланс, половые гормоны, взаимодействующие со стресс-системами и объясняющие половые различия в реактивности на стресс, гормоны щитовидной железы, влияющие на метаболизм и энергетику, и многие другие. Однако для первого знакомства с молекулярным уровнем стресса достаточно ясного понимания трёх главных действующих лиц — кортизола, адреналина и норадреналина, — поскольку именно они определяют основную канву стресс-ответа, на фоне которой разворачивается действие всех остальных молекулярных участников.
1.3 Рецепторы как «замки и ключи»
Молекулы гормонов стресса, какими бы важными они ни были, сами по себе не обладают биологической активностью в том смысле, что они не могут напрямую изменить работу клетки. Для того чтобы сигнал был воспринят и преобразован в клеточный ответ, необходимо специфическое взаимодействие между сигнальной молекулой и её рецептором — специализированным белком, расположенным на поверхности клетки или внутри неё. Это взаимодействие традиционно описывается с помощью метафоры «замка и ключа», впервые предложенной немецким химиком Эмилем Фишером в конце девятнадцатого века применительно к взаимодействию ферментов и субстратов, но оказавшейся исключительно продуктивной для понимания рецепторной сигнализации. Подобно тому как ключ определённой формы подходит только к определённому замку, каждый гормон способен связываться только с теми рецепторами, чья структура комплементарна его молекулярной конфигурации. Это обеспечивает специфичность сигнала: несмотря на то что кровь несёт сотни различных сигнальных молекул одновременно, каждая клетка «слышит» только те сообщения, для восприятия которых у неё есть соответствующие рецепторы.
Молекулярная основа специфичности взаимодействия гормона и рецептора определяется трёхмерной структурой обеих молекул и характером химических взаимодействий между ними. Связывание гормона с рецептором происходит благодаря множеству нековалентных взаимодействий: водородных связей, электростатических взаимодействий между заряженными группами, гидрофобных взаимодействий между неполярными участками молекул, а также ван-дер-ваальсовых сил. Каждое отдельное взаимодействие относительно слабо, но их совокупность обеспечивает высокую аффинность — прочность связывания — и одновременно обратимость: гормон может отсоединиться от рецептора, когда его концентрация снижается. Высокая специфичность означает, что даже небольшие изменения в структуре молекулы могут полностью нарушить связывание, что используется в фармакологии для создания агонистов, активирующих рецептор подобно естественному гормону, и антагонистов, блокирующих рецептор и предотвращающих его активацию. Понимание структуры взаимодействия «гормон-рецептор» на атомном уровне стало возможным благодаря методам рентгеноструктурного анализа и криоэлектронной микроскопии и открыло путь к рациональному дизайну лекарственных препаратов.
Связывание гормона с рецептором не является самоцелью — оно служит триггером для изменений в структуре рецепторного белка, которые инициируют внутриклеточные сигнальные события. Это изменение структуры называется конформационной перестройкой: при связывании лиганда (так называют любую молекулу, связывающуюся с рецептором) белок рецептора слегка меняет свою трёхмерную форму, и это изменение передаётся на другие части белка или на белки-партнёры, с которыми рецептор взаимодействует. Можно сказать, что рецептор «переключается» из неактивного состояния в активное, подобно тому как поворот ключа в замке переводит запирающий механизм из одного положения в другое. Характер последующих внутриклеточных событий зависит от типа рецептора: одни рецепторы сами обладают ферментативной активностью, другие связаны с ионными каналами, третьи взаимодействуют с внутриклеточными сигнальными белками. Таким образом, внеклеточный сигнал — присутствие гормона — преобразуется во внутриклеточный сигнал — изменение активности определённых белков внутри клетки.
Рецепторы гормонов стресса принадлежат к нескольким структурно и функционально различным семействам, что объясняет разнообразие временных масштабов и характера клеточных ответов на различные гормоны. Адреналин и норадреналин связываются с адренорецепторами, принадлежащими к обширному семейству рецепторов, сопряжённых с G-белками. Эти рецепторы представляют собой белки, семикратно пронизывающие клеточную мембрану; их внеклеточная часть связывает гормон, а внутриклеточная часть взаимодействует с G-белками — молекулярными «посредниками», передающими сигнал далее к эффекторным ферментам. Активация адренорецепторов происходит в течение секунд и вызывает быстрые изменения в клеточной физиологии. Рецепторы кортизола, напротив, относятся к семейству ядерных рецепторов — белков, расположенных внутри клетки, куда стероидный гормон проникает благодаря своей липофильности. Связывание кортизола с ядерным рецептором превращает последний в фактор транскрипции, способный связываться с определёнными участками ДНК и изменять экспрессию генов. Этот механизм объясняет более медленное, но более продолжительное действие кортизола.
Разнообразие рецепторов к одному и тому же гормону позволяет достигать тканевой специфичности ответа, что является элегантным решением проблемы координации разнородных эффектов. Адреналин, например, взаимодействует с несколькими подтипами адренорецепторов, обозначаемыми греческими буквами альфа и бета с дополнительными цифровыми индексами. Альфа-один-адренорецепторы преобладают в гладкой мускулатуре сосудов кожи и пищеварительного тракта и при активации вызывают сужение этих сосудов. Бета-один-адренорецепторы преобладают в сердце и при активации увеличивают частоту и силу сердечных сокращений. Бета-два-адренорецепторы преобладают в бронхах и сосудах скелетных мышц и при активации вызывают расширение этих структур. Таким образом, одна молекула адреналина, распространяясь с кровью по всему организму, вызывает согласованный набор изменений: кровь отливает от кожи и внутренних органов, направляясь к мышцам, сердце работает мощнее, бронхи расширяются для лучшего снабжения кислородом. Эта координация достигается не за счёт различных сигнальных молекул, а за счёт тканеспецифического распределения различных рецепторов.
Подобная логика применима и к рецепторам кортизола, где существует два основных типа с различной аффинностью и функцией. Минералокортикоидный рецептор обладает высокой аффинностью к кортизолу и занят гормоном уже при базальных, низких его концентрациях; он опосредует тонические, регуляторные функции кортизола в обычных условиях. Глюкокортикоидный рецептор имеет более низкую аффинность и активируется преимущественно при повышенных концентрациях кортизола, характерных для стресса; именно через него реализуются многие эффекты стрессового ответа, включая метаболическую мобилизацию и иммуномодуляцию. Различное соотношение этих двух типов рецепторов в разных тканях и разных отделах мозга определяет специфику ответа на кортизол. Особенно важно соотношение этих рецепторов в гиппокампе, где они участвуют в регуляции обратной связи оси гипоталамус-гипофиз-надпочечники, и нарушение этого баланса связано с рядом стресс-ассоциированных патологий, включая депрессию. Концепция рецепторного дуализма имеет фундаментальное значение для понимания того, почему нормальные колебания кортизола адаптивны, тогда как хронически повышенный уровень становится патогенным.
Количество рецепторов на клетке не является статичным параметром, но динамически регулируется в зависимости от уровня гормона и других факторов, что представляет собой важный механизм модуляции чувствительности клеток к сигналам. При длительном воздействии высоких концентраций гормона клетки часто уменьшают количество соответствующих рецепторов на своей поверхности — процесс, называемый даунрегуляцией. Это можно рассматривать как защитный механизм, предотвращающий чрезмерную стимуляцию, однако при хроническом стрессе даунрегуляция рецепторов может приводить к снижению чувствительности тканей к регуляторным сигналам. Например, хронически повышенный кортизол может вызывать даунрегуляцию глюкокортикоидных рецепторов в гипоталамусе и гипофизе, что нарушает механизм отрицательной обратной связи и способствует ещё большему повышению кортизола — формируется порочный круг. Противоположный процесс — апрегуляция, увеличение числа рецепторов при низких уровнях гормона — также наблюдается и может приводить к гиперчувствительности. Динамическая регуляция рецепторов представляет собой важный уровень пластичности, позволяющий клеткам адаптировать свою чувствительность к меняющимся условиям.
Понимание рецепторной сигнализации имеет прямые следствия для клинической практики и объясняет механизмы действия многих лекарственных препаратов, применяемых при стресс-связанных состояниях. Бета-адреноблокаторы, такие как пропранолол, конкурентно связываются с бета-адренорецепторами, не активируя их, и тем самым блокируют действие адреналина и норадреналина на сердце. Это объясняет их эффективность при тахикардии, гипертонии и некоторых тревожных состояниях, особенно при ситуативной тревоге, связанной с выступлениями. Глюкокортикоидные препараты — синтетические аналоги кортизола, такие как преднизолон и дексаметазон, — активируют глюкокортикоидные рецепторы и используются для подавления воспаления и иммунных реакций, хотя их длительное применение сопряжено с серьёзными побочными эффектами, воспроизводящими последствия хронического стресса. Разработка новых препаратов часто направлена на создание молекул с более избирательным действием на определённые подтипы рецепторов, что позволяет достигать желаемого терапевтического эффекта при минимизации побочных явлений. Концепция «замка и ключа» в этом контексте превращается в практический инструмент рационального дизайна лекарств.
Современные исследования выявляют всё более сложную картину рецепторной сигнализации, выходящую за пределы простой модели «один гормон — один рецептор — один эффект». Рецепторы могут объединяться в димеры и более крупные комплексы, причём гетеродимеры, состоящие из разных типов рецепторов, могут обладать свойствами, отличными от свойств гомодимеров. Один и тот же рецептор может активировать различные внутриклеточные сигнальные пути в зависимости от того, какой лиганд с ним связан — феномен, называемый функциональной селективностью или смещённым агонизмом. Рецепторы взаимодействуют с множеством внутриклеточных белков-партнёров, формируя сигнальные комплексы, состав которых может различаться в разных клетках. Всё это означает, что клеточный ответ на гормон определяется не только присутствием или отсутствием соответствующего рецептора, но и целым контекстом: какие другие рецепторы экспрессированы, какие сигнальные белки доступны, каково функциональное состояние клетки. Эта сложность объясняет, почему предсказание эффектов гормонов и лекарств «в пробирке» не всегда воспроизводится в живом организме и почему одна и та же молекула может оказывать различное действие в различных физиологических и патологических контекстах.
Метафора «замка и ключа», при всей её полезности для первоначального понимания рецепторной сигнализации, со временем была дополнена более динамичными концепциями, отражающими гибкость молекулярных взаимодействий. Современная модель индуцированного соответствия предполагает, что и лиганд, и рецептор подстраивают свою конформацию при взаимодействии друг с другом — не жёсткий ключ входит в жёсткий замок, а два гибких партнёра взаимно адаптируются. Ещё более современная концепция конформационной селекции рассматривает рецептор как молекулу, постоянно флуктуирующую между различными конформационными состояниями, и связывание лиганда лишь стабилизирует одно из этих состояний. Эти концептуальные уточнения важны для понимания того, почему частичные агонисты — лиганды, активирующие рецептор не полностью — могут быть терапевтически полезны, и как аллостерические модуляторы — молекулы, связывающиеся с рецептором в ином месте, нежели основной лиганд, — могут тонко настраивать рецепторную функцию. Фармакология стресс-связанных состояний всё активнее использует эти возможности для разработки препаратов с более благоприятным профилем эффективности и безопасности.
1.4 Гены — инструкции, которые читаются по-разному
Представление о генах как о неизменной программе, жёстко детерминирующей свойства организма, господствовавшее в массовом сознании и отчасти в научном сообществе на протяжении значительной части двадцатого века, претерпело радикальную трансформацию в свете открытий молекулярной биологии последних десятилетий. Современное понимание генома существенно сложнее и динамичнее: гены представляют собой не столько императивные команды, сколько инструкции, которые могут читаться с различной интенсивностью, в различных комбинациях и в различных контекстах. Эта концептуальная революция имеет фундаментальное значение для понимания стресса на молекулярном уровне, поскольку одним из ключевых механизмов, через которые стресс оказывает своё влияние на организм, является именно изменение характера считывания генетической информации. Стрессовое воздействие не меняет последовательность нуклеотидов в ДНК — то есть не вызывает мутаций в традиционном понимании, — но оно способно существенно изменить то, какие гены активны, насколько интенсивно они экспрессируются и как долго сохраняется это изменённое состояние. Понимание этих механизмов открывает принципиально новые перспективы как для объяснения индивидуальных различий в реакции на стресс, так и для разработки терапевтических стратегий.
Центральная догма молекулярной биологии описывает поток генетической информации от ДНК через РНК к белку, однако этот поток не является ни автоматическим, ни равномерным для всех генов. Геном человека содержит приблизительно двадцать тысяч генов, кодирующих белки, однако в каждой конкретной клетке в каждый конкретный момент времени активна лишь определённая часть этого репертуара. Нейрон экспрессирует иной набор генов, чем гепатоцит или иммунная клетка, и именно это различие в профиле экспрессии определяет специализацию клеток при идентичности их генетического материала. Более того, профиль экспрессии генов в одной и той же клетке может динамически изменяться в ответ на внешние сигналы, включая гормоны стресса. Когда кортизол связывается со своим ядерным рецептором, образовавшийся комплекс перемещается в ядро клетки и взаимодействует с определёнными участками ДНК, называемыми глюкокортикоид-респонсивными элементами, усиливая или подавляя транскрипцию прилежащих генов. Таким образом, сигнал стресса, воспринятый на поверхности или внутри клетки, транслируется в изменение генетической программы.
Процесс активации или подавления экспрессии гена представляет собой многоэтапный и тонко регулируемый механизм, включающий множество молекулярных участников. Первым этапом является транскрипция — синтез молекулы информационной РНК на матрице ДНК, осуществляемый ферментом РНК-полимеразой. Для начала транскрипции необходимо, чтобы регуляторная область гена, называемая промотором, была доступна для связывания транскрипционного аппарата, что зависит от состояния хроматина — комплекса ДНК с белками-гистонами. Факторы транскрипции, включая активированные рецепторы гормонов, связываются с ДНК и рекрутируют коактиваторы или корепрессоры, которые модифицируют хроматин и регулируют активность РНК-полимеразы. После синтеза информационная РНК подвергается процессингу, экспортируется из ядра в цитоплазму и служит матрицей для синтеза белка на рибосомах. Каждый из этих этапов может регулироваться, и стресс-индуцированные сигнальные пути воздействуют на множество точек этого процесса. Результатом является изменение концентрации определённых белков в клетке, что и составляет молекулярную основу клеточного ответа на стресс.
Масштаб генетического ответа на стресс поражает воображение: современные методы транскриптомики, позволяющие одновременно оценивать экспрессию тысяч генов, демонстрируют, что острый стресс изменяет активность сотен генов в различных тканях. Исследования на модельных животных и с использованием образцов крови человека выявили характерные транскрипционные «подписи» стресса, включающие активацию генов, связанных с метаболизмом глюкозы, воспалением, ремоделированием тканей, и подавление генов, связанных с ростом, репродукцией, иммунной защитой от патогенов. Эти изменения развиваются во времени: некоторые гены реагируют в течение минут, представляя «ранний ответ», другие — в течение часов или даже дней. Примечательно, что транскрипционный ответ на стресс не идентичен у всех индивидуумов: генетические варианты в регуляторных областях генов, индивидуальные различия в уровне рецепторов, предшествующий опыт стресса — всё это модулирует характер генетического ответа. Эта вариабельность частично объясняет, почему одни люди более уязвимы к последствиям стресса, тогда как другие демонстрируют устойчивость, и открывает перспективы для персонализированного подхода к профилактике и терапии.
Временная динамика изменений экспрессии генов при стрессе имеет критическое значение для понимания различий между адаптивным ответом и патологическими последствиями. При остром стрессе транскрипционные изменения носят преходящий характер: после устранения стрессора и снижения концентрации гормонов стресса экспрессия генов постепенно возвращается к исходному уровню. Однако при хроническом или повторяющемся стрессе картина существенно усложняется. Некоторые гены остаются аномально активированными или подавленными даже после нормализации гормонального фона, что свидетельствует о формировании новых устойчивых состояний регуляции. Особенно важны изменения в экспрессии генов, кодирующих компоненты самих стресс-систем: рецепторы гормонов, ферменты синтеза и деградации нейромедиаторов, белки обратной связи. Изменение экспрессии этих генов может приводить к долгосрочной перестройке реактивности стресс-систем, что лежит в основе феноменов сенситизации — повышенной чувствительности к последующим стрессорам — или, напротив, притупления реакции.
Концепция «генов раннего ответа» и «генов позднего ответа» отражает временну́ю организацию транскрипционного каскада при стрессе. Гены раннего ответа, или так называемые «немедленно-ранние гены», активируются в течение минут после стрессового воздействия и не требуют синтеза новых белков для своей активации — сигнальные каскады напрямую активируют уже присутствующие в клетке факторы транскрипции. К этой категории относятся многие гены, кодирующие сами факторы транскрипции, такие как продукты генов семейства c-fos и c-jun. Эти ранние продукты, в свою очередь, регулируют экспрессию генов «позднего ответа», кодирующих эффекторные белки — ферменты, структурные белки, рецепторы. Такая каскадная организация обеспечивает амплификацию и диверсификацию сигнала: один первичный стимул через последовательные волны транскрипции приводит к изменению экспрессии сотен генов. Изучение этих каскадов важно не только для фундаментального понимания, но и для идентификации потенциальных терапевтических мишеней: воздействие на ранние этапы каскада может предотвратить развитие всего патологического ответа.
Индивидуальные различия в генетическом ответе на стресс отчасти определяются наследственными вариантами в регуляторных областях генов, влияющими на силу и характер транскрипционного ответа. Однонуклеотидные полиморфизмы в промоторах, энхансерах и других регуляторных элементах могут изменять аффинность связывания факторов транскрипции, включая рецепторы глюкокортикоидов, и тем самым модулировать интенсивность ответа данного гена на стресс. Классическим примером является полиморфизм в гене, кодирующем белок FKBP5 — кошаперон глюкокортикоидного рецептора. Определённые варианты этого гена связаны с усиленной индукцией FKBP5 при стрессе, что, в свою очередь, ослабляет функцию глюкокортикоидного рецептора и нарушает механизм обратной связи. Носители таких вариантов демонстрируют повышенный риск развития депрессии и посттравматического стрессового расстройства, особенно при наличии раннего детского стресса. Этот пример иллюстрирует взаимодействие генетической предрасположенности и средовых факторов на молекулярном уровне и показывает, как вариации в «инструкциях по чтению» генов влияют на уязвимость к стрессу.
Важно подчеркнуть, что изменение экспрессии генов при стрессе не является патологией само по себе — это нормальный адаптивный механизм, позволяющий организму перестраивать свою биохимию в соответствии с требованиями среды. Проблемы возникают тогда, когда этот механизм функционирует избыточно, недостаточно или несвоевременно, либо когда адаптивные изменения становятся хроническими и начинают причинять вред. Понимание того, что гены — не фиксированная судьба, а динамически регулируемый ресурс, имеет глубокие философские и практические следствия. С философской точки зрения, это преодолевает жёсткий генетический детерминизм и открывает пространство для понимания того, как опыт и среда участвуют в формировании биологии организма. С практической точки зрения, это означает, что терапевтические вмешательства — будь то фармакологические, психотерапевтические или связанные с образом жизни — могут оказывать своё благотворное действие отчасти через нормализацию паттернов генной экспрессии. Исследования действительно показывают, что успешная психотерапия и антидепрессанты изменяют экспрессию стресс-связанных генов, что подтверждает молекулярную реальность психологических изменений.
Современные технологии секвенирования и биоинформатики позволяют изучать транскрипционный ответ на стресс с беспрецедентной полнотой и разрешением, открывая новые горизонты понимания. Метод секвенирования РНК даёт возможность не только количественно оценить экспрессию каждого гена, но и выявить альтернативные варианты сплайсинга, новые транскрипты, некодирующие РНК с регуляторными функциями. Секвенирование единичных клеток позволяет увидеть гетерогенность транскрипционного ответа внутри ткани и идентифицировать клеточные субпопуляции с различной реакцией на стресс. Пространственная транскриптомика сохраняет информацию о локализации клеток в ткани, что особенно важно для понимания стресс-ответа в мозге с его сложной анатомической организацией. Эти методы генерируют огромные массивы данных, анализ которых требует новых вычислительных подходов и порождает новые вопросы о механизмах координации генной экспрессии в масштабах организма. Интеграция транскриптомных данных с другими уровнями анализа — эпигенетическим, протеомным, метаболомным — представляет собой одну из главных задач современной системной биологии стресса.
1.5 Эпигенетика — молекулярная память стресса
Если изменения экспрессии генов представляют собой механизм немедленной адаптации к стрессу, то эпигенетические модификации обеспечивают более долгосрочную форму молекулярной памяти, позволяющую опыту стресса оставлять устойчивый след в биологии организма. Термин «эпигенетика», буквально означающий «над генетикой» или «поверх генетики», относится к наследуемым изменениям в функции генов, которые не связаны с изменением последовательности ДНК. Эпигенетические механизмы определяют, какие гены доступны для транскрипции в данной клетке, создавая устойчивые паттерны экспрессии, которые могут сохраняться на протяжении клеточных делений и даже передаваться следующим поколениям. Открытие того, что стресс способен индуцировать эпигенетические изменения, произвело революцию в понимании биологических механизмов долгосрочных последствий стресса и предоставило молекулярное объяснение тому, как ранний опыт может влиять на здоровье и поведение на протяжении всей жизни. Эта область исследований также подняла глубокие вопросы о границах между наследственностью и средой, между биологией и биографией.
Основные эпигенетические механизмы включают метилирование ДНК, модификации гистонов и действие некодирующих РНК, каждый из которых вносит свой вклад в регуляцию доступности генов. Метилирование ДНК представляет собой присоединение метильной группы к цитозину — одному из четырёх оснований ДНК — преимущественно в контексте динуклеотида цитозин-гуанин. Метилирование промоторных областей генов, как правило, ассоциировано с подавлением транскрипции, поскольку метильные группы препятствуют связыванию факторов транскрипции и привлекают белки, конденсирующие хроматин. Модификации гистонов — белков, вокруг которых намотана нить ДНК, — включают ацетилирование, метилирование, фосфорилирование и множество других химических модификаций специфических аминокислотных остатков. Эти модификации влияют на плотность упаковки хроматина и на привлечение регуляторных белков, создавая так называемый «гистоновый код», который определяет транскрипционный потенциал участка генома. Некодирующие РНК, включая микроРНК и длинные некодирующие РНК, регулируют экспрессию генов на посттранскрипционном уровне и участвуют в формировании эпигенетических состояний.
Исследования на животных моделях предоставили убедительные доказательства того, что стресс индуцирует эпигенетические изменения, имеющие функциональные последствия для поведения и физиологии. Классические эксперименты группы Майкла Мини на крысах продемонстрировали, что качество материнской заботы в первые недели жизни влияет на метилирование промотора гена глюкокортикоидного рецептора в гиппокампе потомства. Детёныши матерей, демонстрировавших высокий уровень заботы, имели меньше метилирования в этом регионе, больше экспрессию рецептора, более эффективную обратную связь в регуляции кортизола и более низкую тревожность во взрослом возрасте. Детёныши матерей с низким уровнем заботы демонстрировали противоположный профиль. Примечательно, что эти эффекты определялись не генетикой, а опытом: перекрёстное усыновление — когда детёныши воспитывались неродными матерями — показало, что именно поведение приёмной матери, а не биологическое происхождение определяло эпигенетический профиль и поведенческий фенотип потомства. Эти эксперименты стали парадигматическими для всей области эпигенетики стресса.
Перенос этих находок на человека осложнён очевидными этическими ограничениями экспериментальных манипуляций, однако множество исследований указывает на сходные механизмы у людей. Анализ посмертных образцов мозга людей, совершивших суицид и имевших историю детского насилия, выявил повышенное метилирование промотора гена глюкокортикоидного рецептора в гиппокампе — паттерн, аналогичный наблюдаемому у крыс с низким уровнем материнской заботы. Исследования периферических тканей, преимущественно крови, показывают, что неблагоприятный детский опыт ассоциирован с характерными эпигенетическими изменениями, сохраняющимися во взрослом возрасте. Продольные исследования демонстрируют, что хронический стресс у взрослых также индуцирует эпигенетические изменения, которые коррелируют с симптомами депрессии и тревоги. Вместе с тем интерпретация этих данных требует осторожности: большинство исследований на людях носит корреляционный характер, и каузальные связи между эпигенетическими изменениями и клиническими исходами ещё предстоит окончательно установить.
Временна́я стабильность эпигенетических меток существенно варьирует и определяет, какие изменения являются преходящими адаптациями, а какие — устойчивыми следами опыта. Метилирование ДНК традиционно рассматривалось как относительно стабильная модификация, хотя открытие ферментов активного деметилирования показало, что даже эта метка может быть динамичной. Модификации гистонов, как правило, более лабильны и могут изменяться в масштабах минут и часов в ответ на сигналы среды. Однако некоторые комбинации гистоновых модификаций формируют устойчивые состояния хроматина, сохраняющиеся на протяжении клеточных делений благодаря механизмам эпигенетического наследования. Понимание факторов, определяющих стабильность эпигенетических изменений, критически важно для оценки долгосрочных последствий стресса. Парадоксально, но та же пластичность, которая делает эпигеном уязвимым к негативным воздействиям, открывает возможности для терапевтического вмешательства: если эпигенетические изменения обратимы, то целенаправленные воздействия могут потенциально «стирать» патологические метки стресса.
Одним из наиболее интригующих и дискуссионных аспектов эпигенетики стресса является возможность межпоколенческой передачи приобретённых эпигенетических изменений. Исследования на грызунах показали, что стресс, переживаемый родителями до зачатия, может влиять на эпигенетический профиль и поведение потомства, не имевшего непосредственного контакта со стрессором. Эти эффекты обнаруживаются как через материнскую, так и через отцовскую линию, хотя механизмы различаются. Передача через материнскую линию может осуществляться через эпигенетические метки в ооците, а также через внутриутробную среду и постнатальное поведение матери. Передача через отцовскую линию, исключающая эти факторы, предполагает эпигенетическую информацию в сперматозоидах — через метилирование ДНК, модификации гистонов или малые РНК. Некоторые исследования обнаружили эффекты даже во втором и третьем поколениях, что свидетельствует о подлинной трансгенерационной передаче, а не только о межпоколенческих эффектах. Эти данные ставят под вопрос строгое разграничение наследственного и приобретённого и имеют потенциально важные импликации для понимания передачи уязвимости к стресс-связанным расстройствам в семьях.
Эпигенетическая регуляция ключевых генов стресс-системы представляет особый интерес как потенциальный механизм долгосрочной калибровки стресс-реактивности. Помимо упомянутого гена глюкокортикоидного рецептора, эпигенетической регуляции подвержены гены кортиколиберина, рецепторов серотонина, нейротрофического фактора мозга, множества нейропептидов и их рецепторов. Координированные эпигенетические изменения в этих генах могут перепрограммировать всю систему стресс-ответа, делая её более или менее реактивной, более или менее способной к саморегуляции. Концепция «биологического встраивания» описывает, как ранний опыт «встраивается» в биологию через эпигенетические механизмы, формируя траектории развития, которые влияют на здоровье на протяжении всей жизни. Эта концепция объясняет хорошо документированную связь между неблагоприятным детским опытом и повышенным риском разнообразных заболеваний во взрослом возрасте — от психических расстройств до сердечно-сосудистых заболеваний и метаболического синдрома.
Вопрос об обратимости стресс-индуцированных эпигенетических изменений имеет принципиальное значение для терапевтических перспектив. Исследования на животных показывают, что некоторые эпигенетические изменения действительно обратимы при определённых условиях. Обогащённая среда, социальная поддержка, физическая активность способны частично нормализовать эпигенетический профиль у животных, подвергшихся раннему стрессу. Фармакологические агенты, влияющие на эпигенетические механизмы — ингибиторы гистоновых деацетилаз, модуляторы метилирования ДНК, — демонстрируют эффекты в моделях стресс-связанных расстройств, хотя их клиническое применение осложнено низкой специфичностью и потенциальными побочными эффектами. Интересно, что традиционные антидепрессанты и психотерапия также могут оказывать эпигенетические эффекты, нормализуя паттерны метилирования и гистоновых модификаций в определённых генах. Это открывает перспективу «эпигенетической терапии» не как отдельного подхода, а как понимания молекулярных механизмов уже существующих эффективных вмешательств.
Эпигенетика стресса поднимает глубокие вопросы о природе биологической памяти и её отношении к психологической памяти и идентичности. Эпигенетические метки представляют собой форму клеточной памяти, позволяющую клетке «помнить» прошлые воздействия и соответствующим образом настраивать свою функцию. Эта память распределена по миллиардам клеток организма и не локализована в каком-либо одном органе. Она не сознательна и не извлекаема в форме воспоминаний, однако она формирует физиологический контекст, в котором разворачивается сознательный опыт, и влияет на эмоциональные и когнитивные реакции. Можно сказать, что тело «помнит» стресс на молекулярном уровне даже тогда, когда сознательные воспоминания недоступны или отсутствуют. Это особенно релевантно для понимания последствий раннего травматического опыта, который может не сохраняться в эксплицитной памяти, но оставляет эпигенетический след, влияющий на физиологию и поведение. Интеграция эпигенетического и психологического уровней анализа представляет собой одну из захватывающих задач современной науки о стрессе.
1.6 Молекулярные каскады — цепная реакция внутри клетки
Между связыванием гормона стресса с рецептором на поверхности или внутри клетки и конечным изменением клеточной функции разворачивается сложнейшая последовательность молекулярных событий, которую принято называть сигнальным каскадом или путём передачи сигнала. Эти внутриклеточные каскады обеспечивают преобразование внешнего сигнала — присутствия гормона — во внутриклеточный ответ, будь то изменение метаболизма, секреции, сократительной активности или экспрессии генов. Каскадная организация сигнальных путей не является случайностью эволюции, но представляет собой элегантное решение нескольких принципиальных задач: амплификации сигнала, позволяющей небольшому количеству гормона вызывать масштабный ответ; интеграции множественных сигналов на различных уровнях каскада; тонкой регуляции интенсивности и продолжительности ответа; обеспечения специфичности ответа в различных типах клеток. Понимание архитектуры сигнальных каскадов необходимо для объяснения того, как молекулярные события транслируются в физиологические изменения, и для идентификации потенциальных точек терапевтического вмешательства при стресс-связанных нарушениях.
Принцип амплификации сигнала является одной из ключевых характеристик каскадной организации и объясняет, почему пикомолярные и наномолярные концентрации гормонов способны вызывать драматические изменения в работе целого организма. Рассмотрим классический пример адренергической сигнализации: одна молекула адреналина, связавшись с бета-адренорецептором, активирует его; активированный рецептор, в свою очередь, активирует несколько молекул G-белка, с которыми он способен последовательно взаимодействовать за время своего активного состояния. Каждый активированный G-белок активирует молекулу фермента аденилатциклазы, которая катализирует синтез множества молекул циклического аденозинмонофосфата — вторичного посредника. Циклический аденозинмонофосфат активирует протеинкиназу А, каждая молекула которой фосфорилирует множество молекул субстрата. Если субстратом является другой фермент, его активация приводит к преобразованию тысяч молекул метаболита. Таким образом, на каждом этапе каскада происходит многократное усиление: одна молекула гормона в конечном счёте приводит к образованию или преобразованию миллионов молекул внутриклеточных метаболитов.
Временна́я динамика сигнальных каскадов определяет скорость развития клеточного ответа и его продолжительность. Различные каскады характеризуются различными временными масштабами, что соответствует их функциональной роли. Каскады, регулирующие быстрые физиологические ответы — такие как сокращение мышц или секреция, — включают небольшое число этапов и реализуются в течение миллисекунд или секунд. Каскады, ведущие к изменению экспрессии генов, более протяжённы во времени и включают транслокацию сигнальных молекул в ядро, связывание с ДНК, транскрипцию, процессинг РНК, трансляцию и созревание белка — процесс, занимающий от десятков минут до часов. Продолжительность сигнала определяется балансом между активацией и инактивацией на каждом этапе каскада: ферменты деградации вторичных посредников, фосфатазы, удаляющие фосфатные группы с белков, убиквитинлигазы, маркирующие белки для деградации, — все эти механизмы обеспечивают завершение сигнала после устранения первичного стимула. Нарушение этих механизмов выключения может приводить к хронической активации каскадов даже в отсутствие внешнего сигнала, что лежит в основе ряда патологических состояний.
Основные сигнальные каскады, задействованные в стресс-ответе, включают несколько хорошо охарактеризованных путей, каждый из которых имеет свой профиль активации и спектр эффектов. Путь циклического аденозинмонофосфата и протеинкиназы А, упомянутый выше, опосредует многие эффекты катехоламинов и ряда других гормонов, действующих через рецепторы, сопряжённые с G-белками стимулирующего типа. Путь фосфолипазы С и протеинкиназы С активируется через G-белки другого типа и ведёт к мобилизации внутриклеточного кальция и активации множества кальций-зависимых процессов. Митоген-активируемые протеинкиназные пути представляют собой семейство каскадов, передающих сигналы от поверхностных рецепторов к ядру и регулирующих пролиферацию, дифференцировку и выживание клеток. Путь ядерного фактора каппа В является центральным регулятором воспалительного ответа и активируется при различных формах клеточного стресса. Путь глюкокортикоидного рецептора, описанный ранее, действует через прямую регуляцию транскрипции. Эти пути не изолированы, но интенсивно взаимодействуют друг с другом, формируя сложную сигнальную сеть.
Перекрёстное взаимодействие между сигнальными путями, или кросс-ток, представляет собой фундаментальный принцип организации клеточной сигнализации, обеспечивающий интеграцию множественных входящих сигналов. На практике клетка никогда не получает один изолированный сигнал — она одновременно воспринимает десятки гормонов, нейромедиаторов, цитокинов и других сигнальных молекул, и её ответ определяется интеграцией всей этой информации. Кросс-ток осуществляется через несколько механизмов: общие компоненты различных каскадов, которые служат точками конвергенции; регуляция компонентов одного каскада продуктами другого; конкуренция за общие эффекторы. Например, глюкокортикоидный рецептор и ядерный фактор каппа В взаимно ингибируют друг друга на уровне транскрипции, что объясняет противовоспалительное действие кортизола: активация глюкокортикоидного рецептора подавляет транскрипционную активность ядерного фактора каппа В и, следовательно, экспрессию провоспалительных генов. Такого рода антагонистические и синергические взаимодействия между каскадами создают сложную логику клеточного ответа, далеко выходящую за рамки простой линейной причинности.
Пространственная организация сигнальных каскадов внутри клетки обеспечивает дополнительный уровень регуляции специфичности и эффективности передачи сигнала. Компоненты каскадов не свободно диффундируют в цитоплазме, но организованы в комплексы и локализованы в определённых субклеточных компартментах. Якорные белки удерживают ферменты и их субстраты в непосредственной близости друг к другу, повышая эффективность передачи сигнала и предотвращая неспецифическую активацию. Мембранные микродомены концентрируют рецепторы и сигнальные молекулы, создавая сигнальные платформы. Транслокация сигнальных молекул между компартментами — например, из цитоплазмы в ядро — представляет собой регулируемый этап каскада и точку контроля. Эта пространственная организация объясняет, как клетка может использовать один и тот же вторичный посредник — например, циклический аденозинмонофосфат — для регуляции различных процессов в различных её частях: локальное повышение концентрации вблизи определённых эффекторов вызывает специфический ответ без влияния на другие потенциальные мишени.
Каскадная организация, при всех её преимуществах, создаёт множество точек потенциальной уязвимости, где нарушения могут приводить к патологическим последствиям. Мутации в генах, кодирующих компоненты каскадов, могут вызывать их конститутивную активацию или, напротив, инактивацию, что лежит в основе ряда заболеваний. Хроническая стимуляция каскадов при длительном воздействии гормонов стресса может приводить к истощению компонентов, десенситизации или, напротив, к патологическому усилению отдельных ветвей. Нарушения механизмов выключения сигнала — дефицит фосфатаз, ферментов деградации вторичных посредников — могут вызывать персистирующую активацию даже после устранения первичного стимула. При хроническом стрессе эти нарушения каскадной регуляции могут кумулироваться и вносить вклад в развитие патологических состояний. Понимание конкретных молекулярных поломок в сигнальных каскадах открывает возможности для таргетной терапии, направленной на нормализацию сигнализации на определённом уровне каскада.
Фармакологическое воздействие на сигнальные каскады представляет собой один из основных механизмов действия лекарственных препаратов, включая средства для лечения стресс-связанных расстройств. Многие психотропные препараты прямо или опосредованно влияют на внутриклеточную сигнализацию. Литий, используемый при биполярном расстройстве, ингибирует ряд ферментов каскадов, включая инозитолмонофосфатазу и киназу гликогенсинтазы третьего типа. Антидепрессанты, повышающие уровень моноаминов в синапсах, в конечном счёте реализуют свои эффекты через активацию внутриклеточных каскадов и изменение экспрессии генов в постсинаптических нейронах. Разработка новых препаратов всё чаще ориентируется на конкретные компоненты каскадов — киназы, фосфатазы, адаптерные белки, — что позволяет достигать более специфических эффектов. Однако сложность и взаимосвязанность сигнальных сетей означает, что вмешательство в одну точку неизбежно затрагивает множество связанных процессов, что объясняет профиль побочных эффектов многих препаратов и сложность предсказания эффектов новых соединений.
Системный взгляд на сигнальные каскады признаёт, что они функционируют не как изолированные линейные пути, но как элементы сложной сигнальной сети с многочисленными петлями обратной связи, точками ветвления и конвергенции. Современные методы системной биологии позволяют картировать эти сети и моделировать их динамику с использованием математических подходов. Такие модели выявляют эмерджентные свойства сигнальных сетей, которые не очевидны из анализа отдельных компонентов: бистабильность, позволяющую клетке переключаться между дискретными состояниями; осцилляции, обеспечивающие периодическую модуляцию активности; адаптацию, позволяющую клетке реагировать на изменения, а не на абсолютный уровень сигнала. Применительно к стрессу сетевой подход позволяет понять, как возникают патологические состояния — как точки устойчивого равновесия сети смещаются под влиянием хронического стресса, и как терапевтические вмешательства могут возвращать систему к адаптивному режиму функционирования. Интеграция знаний о сигнальных каскадах с пониманием более высоких уровней организации — клеточного, системного, организменного — составляет одну из центральных задач современной науки о стрессе.
2. Клеточный уровень — базовые единицы стресс-ответа
2.1 Клетка как самостоятельная «фабрика» стресс-ответа
Традиционное представление о клетке как о простейшем «кирпичике» организма, пассивно выполняющем команды, поступающие от нервной и эндокринной систем, не выдерживает критического анализа в свете современных данных клеточной биологии. Каждая из приблизительно тридцати семи триллионов клеток человеческого тела представляет собой самостоятельную биологическую систему поразительной сложности, способную воспринимать сигналы из внешней и внутренней среды, обрабатывать эту информацию и формировать адаптивный ответ задолго до того, как организменные регуляторные системы «узнают» о возникшей проблеме. Более того, концепция клеточного стресса, или стресса на уровне отдельной клетки, предшествует концепции организменного стресса в эволюционном отношении: одноклеточные организмы вынуждены были развить механизмы адаптации к неблагоприятным условиям задолго до появления многоклеточности с её специализированными регуляторными системами. Эти древние механизмы клеточного стресс-ответа сохранились в клетках сложных многоклеточных организмов и составляют фундаментальный уровень, на котором разворачиваются все более высокоорганизованные формы адаптации. Понимание клетки как автономной «фабрики» стресс-ответа необходимо для осознания того, что системные проявления стресса — гормональные сдвиги, изменения поведения, психологические переживания — в конечном счёте реализуются через совокупность клеточных событий и зависят от состояния и компетентности отдельных клеток.
Внутренняя организация клетки может быть продуктивно осмыслена через метафору высокотехнологичного производственного предприятия, в котором различные подразделения выполняют специализированные функции при непрерывной координации их деятельности. Ядро клетки функционирует как «диспетчерский центр» и архив генетической информации, содержащий полный набор инструкций для синтеза всех белков, которые клетка способна производить. Рибосомы, расположенные в цитоплазме и на мембранах эндоплазматического ретикулума, представляют собой «производственные линии», где генетическая информация транслируется в белковые молекулы — рабочие инструменты и структурные компоненты клетки. Эндоплазматический ретикулум и аппарат Гольджи обеспечивают «сборку и упаковку» белков, их модификацию и направление к месту назначения — будь то внутриклеточные органеллы, клеточная мембрана или внеклеточное пространство. Митохондрии функционируют как «энергетические станции», преобразующие энергию питательных веществ в универсальную энергетическую валюту клетки — аденозинтрифосфат. Лизосомы и протеасомы составляют «систему утилизации и переработки», разрушая повреждённые или ненужные компоненты и освобождая их составляющие для повторного использования. Эта функциональная организация не является статичной, но представляет собой динамически регулируемую систему, способную быстро перестраиваться в ответ на изменение условий.
При воздействии стрессора на клетку — будь то гормональный сигнал системного стресса, локальное повреждение, дефицит кислорода или питательных веществ, температурный шок или токсическое воздействие — вся внутренняя «экономика» клетки претерпевает координированную перестройку. Эта перестройка направлена на достижение нескольких целей: минимизацию повреждений, устранение возникших нарушений, адаптацию метаболизма к новым условиям и подготовку к возможному усугублению ситуации. На практике это означает, что одни «производства» резко усиливаются, тогда как другие приостанавливаются или полностью останавливаются. Синтез так называемых белков теплового шока — универсальных молекулярных «шаперонов», помогающих другим белкам сохранять правильную структуру или восстанавливать её после повреждения, — многократно усиливается практически при любой форме клеточного стресса. Одновременно синтез «роскошных» белков, необходимых для роста и пролиферации, но не для немедленного выживания, временно подавляется. Энергетический метаболизм перестраивается в направлении мобилизации резервов и повышения эффективности производства энергии. Системы репарации ДНК активируются для устранения повреждений генетического материала. Антиоксидантные системы усиливаются для нейтрализации повреждающих активных форм кислорода. Все эти изменения координируются сложной сетью сигнальных путей, которые интегрируют информацию о состоянии клетки и определяют оптимальную стратегию ответа.
Концепция клеточного стресс-ответа получила экспериментальное обоснование благодаря открытию универсальных механизмов, активирующихся в клетках при неблагоприятных воздействиях независимо от их конкретной природы. Открытие белков теплового шока в лаборатории итальянского генетика Феруччо Ритосса в шестидесятых годах двадцатого века положило начало систематическому изучению этих механизмов. Исследователи обнаружили, что кратковременное повышение температуры вызывает характерный паттерн активации генов в клетках дрозофилы, и продукты этих генов — белки теплового шока — защищают клетку от последующего более сильного теплового воздействия. Впоследствии оказалось, что эти белки синтезируются в ответ на разнообразные стрессоры — не только тепловой шок, но и окислительный стресс, тяжёлые металлы, гипоксию, инфекции, и многие другие повреждающие факторы. Более того, система белков теплового шока оказалась эволюционно консервативной, присутствующей у всех изученных организмов от бактерий до человека, что свидетельствует о её фундаментальной роли в клеточной адаптации. Помимо системы белков теплового шока были идентифицированы другие универсальные программы клеточного стресс-ответа: ответ на нескладчатые белки в эндоплазматическом ретикулуме, ответ на повреждение ДНК, аутофагия — контролируемое «самопоедание» повреждённых компонентов, и ряд других.
Взаимоотношения между клеточным стрессом и системным, организменным стрессом носят сложный двунаправленный характер, понимание которого критически важно для интеграции молекулярного и клеточного уровней анализа. С одной стороны, системный стресс — воспринятая мозгом угроза, активация гормональных осей, выброс кортизола и катехоламинов — достигает индивидуальных клеток в форме гормональных сигналов и индуцирует характерные клеточные ответы. Кортизол, проникая в клетки, связывается с глюкокортикоидным рецептором и изменяет экспрессию тысяч генов, перестраивая метаболизм и функцию клетки в соответствии с требованиями системного стресс-ответа. С другой стороны, первичный клеточный стресс — например, локальное повреждение ткани или инфекция — генерирует сигналы, которые распространяются на системный уровень и могут вызывать полноценный организменный стресс-ответ. Цитокины, выделяемые клетками в зоне повреждения, достигают мозга и активируют гипоталамо-гипофизарно-надпочечниковую ось, вызывая системное повышение кортизола. Эта двунаправленность означает, что клеточный и системный уровни стресса не могут рассматриваться изолированно — они составляют единую интегрированную систему, в которой события на одном уровне неизбежно влияют на другой.
Важнейшей характеристикой клеточного стресс-ответа является его дозозависимость и временна́я динамика, определяющие, будет ли исход адаптивным или патологическим. Умеренный стрессор, не превышающий адаптивного потенциала клетки, вызывает транзиторную активацию защитных механизмов, которые не только устраняют возникшие повреждения, но и могут повысить устойчивость клетки к последующим воздействиям — феномен, известный как гормезис или прекондиционирование. Классическим примером является ишемическое прекондиционирование: кратковременные эпизоды умеренной ишемии защищают сердце от повреждения при последующем инфаркте через активацию клеточных защитных программ. Однако интенсивный или продолжительный стрессор может превысить адаптивные возможности клетки, приводя к накоплению повреждений, истощению защитных ресурсов и в конечном счёте к гибели клетки либо к переходу в дисфункциональное состояние хронического стресса. Критически важно понимать, что между полной адаптацией и гибелью клетки существует целый спектр промежуточных состояний, включая обратимое нарушение функции, переход в состояние покоя, сенесценцию — состояние старения с прекращением делений, но сохранением жизнеспособности. Именно эти промежуточные состояния часто лежат в основе хронических патологических процессов, связанных с продолжительным стрессом.
Энергетический аспект клеточного стресс-ответа заслуживает особого внимания, поскольку адаптация к стрессу является энергозатратным процессом, требующим перераспределения ресурсов от обычных функций к защитным. Синтез белков теплового шока и других стресс-индуцируемых белков потребляет значительное количество аденозинтрифосфата и аминокислот. Репарация повреждённой ДНК, удаление дефектных белков через протеасомную деградацию, аутофагия — все эти процессы энергозависимы. В условиях острого стресса клетка способна мобилизовать резервы и временно увеличить энергопродукцию, покрывая возросшие потребности. Однако при хроническом стрессе энергетический баланс клетки может нарушаться: постоянно высокие требования к энергопродукции истощают резервы, перегружают митохондрии, ведут к накоплению окислительных повреждений самих энергетических станций. Это создаёт порочный круг, в котором снижение энергопродукции ограничивает возможности адаптации, а недостаточная адаптация приводит к дальнейшим повреждениям. Концепция клеточной энергетической недостаточности как основы многих стресс-связанных патологий приобретает всё большее признание и будет детально рассмотрена в контексте митохондриальной функции.
Гетерогенность клеток организма определяет различия в их чувствительности и характере ответа на стресс, что имеет важные следствия для понимания тканеспецифичных последствий стресса. Различные типы клеток экспрессируют различные наборы рецепторов к гормонам стресса, обладают различным уровнем антиоксидантной защиты, различной способностью к репарации и регенерации. Нейроны, будучи постмитотическими клетками с высочайшими энергетическими потребностями и ограниченной способностью к замещению, особенно уязвимы к повреждающему действию хронического стресса. Клетки печени, напротив, обладают мощными системами детоксикации и высоким регенеративным потенциалом, что делает их относительно устойчивыми. Клетки иммунной системы занимают особое положение, будучи одновременно мишенями стрессовых воздействий и активными участниками стресс-ответа, модулирующими реакции других клеток и тканей. Понимание тканеспецифических особенностей клеточного стресс-ответа позволяет объяснить, почему хронический стресс преимущественно поражает определённые органы и системы — мозг, сердечно-сосудистую систему, желудочно-кишечный тракт, — относительно щадя другие. Эта гетерогенность также имеет терапевтические импликации: идеальное вмешательство должно учитывать различия в клеточных ответах и таргетировать наиболее уязвимые популяции.
Осознание клетки как самостоятельной «фабрики» стресс-ответа имеет глубокие концептуальные и практические следствия для всей науки о стрессе. Концептуально это означает, что системный стресс-ответ не является чем-то, что «накладывается» на пассивные клетки сверху, но представляет собой эмерджентный результат координации миллиардов автономных клеточных ответов, интегрируемых нервной и эндокринной системами. Каждая клетка одновременно является и исполнителем команд, поступающих от системных регуляторов, и самостоятельным «агентом», реагирующим на своё локальное окружение и посылающим сигналы обратно к системному уровню. Практически это означает, что терапевтические вмешательства могут таргетировать не только системные механизмы — гормональные оси, нейромедиаторные системы, — но и непосредственно клеточные процессы: поддержание энергетического метаболизма, усиление антиоксидантной защиты, модуляцию аутофагии, нормализацию белкового гомеостаза. Ряд перспективных терапевтических подходов, основанных на понимании клеточного стресс-ответа, включая применение индукторов белков теплового шока, модуляторов аутофагии и митохондриально-таргетных антиоксидантов, находятся на различных стадиях исследования и разработки.
2.2 Нейроны — специалисты по быстрой передаче сигналов
Среди всего многообразия клеточных типов, составляющих человеческий организм, нейроны занимают совершенно особое положение благодаря своей уникальной способности генерировать и проводить электрические сигналы с высокой скоростью и точностью. Эта способность превратила нервную систему в главный интегратор и координатор всех функций организма, включая восприятие стрессоров, организацию стресс-ответа и регуляцию возвращения к гомеостазу. Нейроны отличаются от других клеток организма не только функционально, но и морфологически: их вытянутая форма с длинными отростками — аксонами и дендритами — оптимизирована для приёма, обработки и передачи информации на значительные расстояния. Один мотонейрон спинного мозга может иметь аксон длиной более метра, связывающий его с мышцей конечности; пирамидный нейрон коры головного мозга формирует тысячи синаптических контактов с другими нейронами, интегрируя информацию из множества источников. Эта архитектура делает нейроны незаменимыми для задач быстрой коммуникации, но одновременно создаёт уникальные уязвимости, которые становятся критически важными в контексте стресса. Понимание того, как нейроны функционируют в норме и как стресс изменяет их работу, составляет необходимую основу для всех последующих обсуждений нейробиологии стресса.
Электрическая сигнализация нейронов основана на фундаментальном свойстве клеточной мембраны — способности поддерживать разность электрических потенциалов между внутренней и внешней средой клетки. В состоянии покоя внутренняя сторона мембраны нейрона имеет отрицательный заряд относительно внешней, составляющий примерно минус семьдесят милливольт — так называемый мембранный потенциал покоя. Это состояние поддерживается активностью ионных насосов, непрерывно откачивающих ионы натрия из клетки и закачивающих ионы калия внутрь, а также избирательной проницаемостью мембраны для различных ионов. Когда нейрон получает достаточно сильный стимул — обычно в форме химического сигнала от другого нейрона или сенсорного рецептора, — происходит резкое изменение проницаемости мембраны: открываются потенциал-зависимые натриевые каналы, ионы натрия устремляются внутрь клетки, и мембранный потенциал скачкообразно сдвигается в положительную сторону. Это событие называется потенциалом действия или нервным импульсом и представляет собой основную единицу информации в нервной системе. Потенциал действия распространяется вдоль аксона нейрона подобно волне, достигая нервных окончаний за считаные миллисекунды и обеспечивая передачу сигнала на большие расстояния без затухания.
Скорость проведения нервного импульса по аксону определяется его структурными характеристиками и может варьировать от менее одного метра в секунду до более ста метров в секунду. Наиболее быстрое проведение обеспечивается в миелинизированных волокнах, где аксон окружён оболочкой из миелина — жироподобного вещества, производимого специализированными глиальными клетками. Миелиновая оболочка прерывается в регулярных интервалах — перехватах Ранвье, — и потенциал действия «перескакивает» от одного перехвата к другому, что многократно ускоряет проведение. Эта организация имеет прямое отношение к стрессу: хронический стресс может нарушать целостность миелина, приводя к замедлению проведения и нарушению временно́й координации нейронных сигналов. Современные нейровизуализационные исследования демонстрируют изменения белого вещества мозга — состоящего преимущественно из миелинизированных аксонов — у людей с хроническим стрессом и стресс-связанными расстройствами, что может отражать повреждение или недоразвитие миелиновых оболочек. Понимание структурной уязвимости нейронов к стрессу дополняет картину функциональных изменений.
Синапс — место контакта между нейронами — представляет собой критически важную структуру, в которой электрический сигнал преобразуется в химический, а затем снова в электрический. Когда потенциал действия достигает нервного окончания — пресинаптического терминала, — открываются потенциал-зависимые кальциевые каналы, и входящий кальций запускает слияние синаптических везикул, содержащих нейромедиатор, с пресинаптической мембраной. Нейромедиатор высвобождается в узкую синаптическую щель шириной около двадцати нанометров и диффундирует к постсинаптической мембране, где связывается с рецепторами. В зависимости от типа рецептора и природы нейромедиатора это связывание может вызвать деполяризацию постсинаптической мембраны, приближающую нейрон к порогу генерации потенциала действия — возбуждающий постсинаптический потенциал, — либо гиперполяризацию, удаляющую от порога — тормозный постсинаптический потенциал. Каждый нейрон интегрирует тысячи таких постсинаптических потенциалов от множества синапсов, и только если их сумма превышает определённый порог, генерируется выходной потенциал действия. Эта интегративная функция делает нейрон не просто проводником, но вычислительным элементом, принимающим «решение» о том, передавать ли сигнал далее.
Стресс оказывает многогранное влияние на синаптическую передачу, затрагивая практически все этапы этого процесса. Гормоны стресса модулируют высвобождение нейромедиаторов: норадреналин и кортикотропин-рилизинг гормон усиливают выброс возбуждающих нейромедиаторов в ряде синапсов, повышая общий уровень возбуждения нейронных сетей. Одновременно стресс влияет на обратный захват нейромедиаторов — процесс их удаления из синаптической щели, — что изменяет продолжительность и интенсивность синаптического сигнала. Количество и чувствительность постсинаптических рецепторов также регулируются при стрессе: хроническое воздействие может приводить к даунрегуляции рецепторов, снижающей эффективность синаптической передачи. Структурные изменения синапсов при хроническом стрессе документированы многочисленными исследованиями: в гиппокампе и префронтальной коре наблюдается сокращение числа синапсов и упрощение дендритных деревьев нейронов, тогда как в миндалине, напротив, может происходить увеличение синаптической плотности. Эти структурные перестройки коррелируют с функциональными изменениями — нарушениями памяти, исполнительных функций, эмоциональной регуляции, — которые характерны для хронического стресса.
Электрическая активность нейронов организована не только на уровне индивидуальных потенциалов действия, но и на уровне паттернов, характеризующих частоту и временну́ю структуру «стрельбы» нейрона. Нейрон может генерировать одиночные импульсы, пачки импульсов с определённой частотой, ритмические осцилляции, или паузы между периодами активности — и каждый из этих паттернов несёт различную информацию для постсинаптических нейронов и для регулируемых эффекторов. Стресс существенно изменяет паттерны нейронной активности в ключевых структурах мозга, участвующих в стресс-ответе. В миндалине, детектирующей угрозы, острый стресс вызывает повышение частоты нейронной «стрельбы» и усиление синхронизации между нейронами. В префронтальной коре, обеспечивающей когнитивный контроль и регуляцию эмоций, хронический стресс, напротив, снижает активность и нарушает нормальные паттерны осцилляций. В гиппокампе, критическом для памяти и контекстуальной обработки, стресс подавляет так называемую долговременную потенциацию — клеточный механизм формирования памяти, — и может усиливать долговременную депрессию — механизм ослабления синаптических связей. Эти изменения паттернов активности лежат в основе когнитивных и эмоциональных проявлений стресса.
Нейромедиаторные системы, вовлечённые в стресс-ответ, включают широкий спектр химических посредников с различными функциональными профилями. Глутамат — основной возбуждающий нейромедиатор мозга — играет ключевую роль в быстрой синаптической передаче и в механизмах синаптической пластичности. Однако избыточное высвобождение глутамата при сильном или продолжительном стрессе может вызывать эксайтотоксичность — повреждение и гибель нейронов вследствие чрезмерной активации глутаматных рецепторов и последующего кальциевого перегруза. Гамма-аминомасляная кислота — основной тормозный нейромедиатор — обеспечивает контроль над возбудимостью нейронных сетей и предотвращает чрезмерную активацию. Нарушение баланса между глутаматергическим возбуждением и ГАМКергическим торможением при хроническом стрессе ассоциировано с тревожностью и депрессией. Моноаминовые нейромедиаторы — серотонин, норадреналин, дофамин — модулируют активность обширных областей мозга из относительно небольших ядер ствола и среднего мозга и критически вовлечены в регуляцию настроения, мотивации, внимания. Нейропептиды — кортикотропин-рилизинг гормон, нейропептид Y, окситоцин и многие другие — оказывают модулирующее влияние на нейронные сети и участвуют в формировании сложных поведенческих и эмоциональных ответов на стресс. Понимание этой нейрохимической сложности необходимо для объяснения разнообразия проявлений стресса и механизмов действия психотропных препаратов.
Нейропластичность — способность нейронов и нейронных сетей изменять свою структуру и функцию в ответ на опыт — представляет собой двусторонний меч в контексте стресса. С одной стороны, пластичность обеспечивает адаптацию к меняющимся условиям и лежит в основе обучения и памяти. С другой стороны, та же пластичность означает, что стрессовый опыт может вызывать устойчивые изменения в нейронных цепях, лежащие в основе долгосрочных последствий стресса. Исследования на животных моделях демонстрируют, что хронический стресс вызывает ретракцию дендритов и потерю дендритных шипиков — постсинаптических структур, на которых формируются возбуждающие синапсы, — в пирамидных нейронах гиппокампа и префронтальной коры. Эти структурные изменения коррелируют с нарушениями пространственной памяти и исполнительных функций. Параллельно в миндалине наблюдается противоположная картина — рост дендритов и увеличение синаптической плотности, что может лежать в основе усиленной тревожности. Важно, что эти изменения частично обратимы: прекращение стресса, обогащённая среда, физическая активность, некоторые фармакологические вмешательства способствуют восстановлению нормальной структуры нейронов. Эта обратимость даёт надежду на терапевтическое восстановление после стресса.
Энергетическая уязвимость нейронов составляет один из ключевых факторов их чувствительности к стрессу и заслуживает особого рассмотрения. Мозг, составляя лишь около двух процентов массы тела, потребляет приблизительно двадцать процентов всего кислорода и глюкозы организма, что отражает исключительную метаболическую активность нейронов. Поддержание мембранного потенциала, синаптическая передача, внутриклеточный транспорт вдоль длинных аксонов — все эти процессы требуют непрерывного снабжения энергией в форме аденозинтрифосфата. В отличие от многих других клеток, нейроны практически не располагают запасами энергетических субстратов и критически зависят от непрерывного поступления глюкозы и кислорода с кровотоком. Стресс создаёт дополнительную нагрузку на энергетический метаболизм нейронов: усиленная нейронная активность повышает потребности в энергии, тогда как глюкокортикоиды и катехоламины изменяют метаболизм глюкозы. При хроническом стрессе этот энергетический дефицит может приводить к нарушению функции нейронов и делать их уязвимыми к дополнительным повреждающим воздействиям. Связь между митохондриальной дисфункцией в нейронах и нейропсихиатрическими последствиями хронического стресса активно исследуется в современной нейробиологии.
Интеграция нейронов в функциональные сети переносит анализ стресса с уровня индивидуальных клеток на уровень систем, что будет детально рассмотрено в последующих разделах курса. Однако уже на этапе обсуждения клеточного уровня важно подчеркнуть, что изменения в индивидуальных нейронах приобретают свой функциональный смысл только в контексте их принадлежности к определённым цепям и сетям. Нейрон гиппокампа с редуцированным дендритным деревом получает меньше входящей информации и вносит искажённый вклад в работу гиппокампальной сети, что проявляется в нарушениях памяти. Нейрон миндалины с повышенной возбудимостью усиливает активность сети страха, что проявляется в тревожности. Понимание того, как клеточные изменения транслируются в изменения сетевой активности и далее в поведенческие и психологические феномены, представляет собой одну из центральных задач нейробиологии стресса. Это понимание требует методов, позволяющих одновременно отслеживать активность множества нейронов и связывать её с поведением, — методов, которые стремительно развиваются благодаря достижениям оптогенетики, многоэлектродной регистрации и функциональной нейровизуализации.
2.3 Иммунные клетки — мобильные защитники и сигнальщики
Иммунная система традиционно рассматривалась как автономный защитный механизм, функционирующий относительно независимо от нервной системы и психологических процессов, однако исследования последних десятилетий радикально изменили это представление. Иммунные клетки оказались не только защитниками организма от патогенов, но и важнейшими участниками стресс-ответа, двунаправленно взаимодействующими с нервной системой и вносящими существенный вклад в физиологические и психологические последствия стресса. Понимание этого взаимодействия привело к формированию новой междисциплинарной области — психонейроиммунологии, изучающей связи между психологическими процессами, нервной системой и иммунитетом. Иммунные клетки обладают рецепторами к гормонам стресса и нейромедиаторам, что делает их чувствительными к сигналам нервной системы, и одновременно производят сигнальные молекулы — цитокины, — способные влиять на функцию мозга и формировать поведенческие и эмоциональные проявления. Эта интеграция иммунной и нервной систем имеет глубокий эволюционный смысл: стресс часто ассоциирован с риском травмы и инфекции, и координированная активация защитных механизмов повышает шансы выживания. Однако в условиях современной жизни, где большинство стрессоров не несут непосредственной физической угрозы, эта связь может становиться источником патологических последствий.
Разнообразие иммунных клеток отражает многогранность задач иммунной защиты и определяет различные аспекты их участия в стресс-ответе. Врождённый иммунитет представлен клетками, обеспечивающими быстрый, хотя и относительно неспецифический ответ на угрозы: нейтрофилы являются наиболее многочисленными лейкоцитами крови и первыми прибывают к месту инфекции или повреждения, поглощая и уничтожая патогены; макрофаги, помимо фагоцитоза, играют ключевую роль в презентации антигенов и продукции цитокинов, координирующих воспалительный ответ; натуральные киллеры уничтожают инфицированные вирусами и опухолевые клетки без предварительной сенсибилизации. Адаптивный иммунитет представлен лимфоцитами, обеспечивающими высокоспецифичный ответ на конкретные патогены и формирующими иммунологическую память: Т-лимфоциты включают хелперы, координирующие иммунный ответ, цитотоксические клетки, уничтожающие инфицированные клетки, и регуляторные клетки, контролирующие интенсивность ответа; В-лимфоциты продуцируют антитела, нейтрализующие патогены и маркирующие их для уничтожения. Каждый из этих клеточных типов по-своему реагирует на стресс и вносит специфический вклад в иммунные последствия стрессовых воздействий.
Влияние стресса на иммунные клетки определяется сложным взаимодействием нервных и гормональных сигналов, достигающих иммунной системы через несколько каналов. Симпатическая нервная система иннервирует первичные и вторичные лимфоидные органы — тимус, селезёнку, лимфатические узлы, костный мозг, — и высвобождение норадреналина из нервных окончаний напрямую воздействует на иммунные клетки через адренорецепторы на их поверхности. Гипоталамо-гипофизарно-надпочечниковая ось обеспечивает гормональный путь влияния: кортизол и другие глюкокортикоиды связываются с глюкокортикоидными рецепторами иммунных клеток и оказывают мощное модулирующее действие на их функцию. Катехоламины — адреналин и норадреналин — выделяемые мозговым веществом надпочечников, также достигают иммунных клеток через кровоток. Эти нейроэндокринные сигналы вызывают комплекс изменений в иммунных клетках: изменение экспрессии генов, перераспределение между тканями и кровеносным руслом, модуляцию продукции цитокинов, изменение цитотоксической активности и способности к фагоцитозу. Результат этих изменений существенно зависит от характера стресса — его интенсивности, продолжительности и контекста.
Острый стресс, как правило, оказывает усиливающее действие на ряд параметров иммунной функции, что может быть понято как подготовка к потенциальной травме и инфекции. В первые минуты и часы после воздействия острого стрессора наблюдается перераспределение иммунных клеток: они мобилизуются из депо — селезёнки, костного мозга — в кровоток и далее в ткани, наиболее подверженные риску повреждения, — кожу, слизистые оболочки, лимфатические узлы. Функциональная активность иммунных клеток при остром стрессе также может повышаться: усиливается цитотоксичность натуральных киллеров, способность макрофагов к фагоцитозу, продукция провоспалительных цитокинов. Эти изменения адаптивны в эволюционном контексте: они обеспечивают готовность к быстрому ответу на инфекцию, которая может проникнуть через повреждённые ткани. Классические эксперименты демонстрируют, что животные, подвергнутые острому стрессу непосредственно перед вакцинацией, формируют более сильный иммунный ответ, чем контрольные животные. Однако этот усиливающий эффект ограничен во времени и сменяется иммуносупрессией при продолжении или повторении стресса.
Хронический стресс оказывает качественно иное, преимущественно супрессивное влияние на адаптивный иммунитет, одновременно способствуя развитию хронического воспаления — парадоксальная комбинация, имеющая серьёзные последствия для здоровья. При длительном воздействии стресса наблюдается подавление пролиферации лимфоцитов, снижение продукции антител, ослабление цитотоксической активности, нарушение формирования иммунологической памяти. Это объясняет хорошо документированную связь между хроническим стрессом и повышенной восприимчивостью к инфекциям: люди, переживающие хронический стресс — ухаживающие за больными родственниками, находящиеся в состоянии затяжного конфликта или экономической нестабильности, — демонстрируют более слабый ответ на вакцинацию, более длительное заживление ран, более частые и тяжёлые простудные заболевания. Механизмы этой иммуносупрессии включают прямое подавляющее действие хронически повышенного кортизола на лимфоциты, истощение иммунных ресурсов, нарушение нормального созревания и дифференцировки иммунных клеток. Однако параллельно с подавлением адаптивного иммунитета хронический стресс активирует другую ветвь иммунного ответа — системное воспаление низкой интенсивности.
Системное воспаление при хроническом стрессе представляет собой ключевой механизм, связывающий стресс с широким спектром заболеваний — от сердечно-сосудистой патологии до депрессии и нейродегенерации. В отличие от острого воспаления, направленного на устранение конкретной угрозы и самоограничивающегося во времени, хроническое воспаление низкой интенсивности персистирует без очевидного внешнего триггера и оказывает повреждающее действие на ткани. При хроническом стрессе повышаются уровни провоспалительных цитокинов — интерлейкина-шесть, фактора некроза опухоли альфа, интерлейкина-один-бета — в крови, даже в отсутствие инфекции или явного повреждения. Макрофаги и другие клетки врождённого иммунитета переключаются на провоспалительный фенотип, продуцируя больше воспалительных медиаторов. Одновременно ослабляется функция регуляторных Т-лимфоцитов, которые в норме контролируют интенсивность воспалительного ответа и предотвращают его чрезмерность. Результатом является состояние, которое исследователи описывают метафорой «тлеющего» воспаления — недостаточно интенсивного для острых симптомов, но достаточного для кумулятивного повреждения тканей на протяжении месяцев и лет.
Цитокины — сигнальные белки, продуцируемые иммунными клетками — представляют собой ключевое звено, через которое иммунная система влияет на мозг и формирует психологические проявления стресса. Цитокины способны пересекать гематоэнцефалический барьер или сигнализировать мозгу через специальные транспортные системы и афферентные нервные пути, в частности через блуждающий нерв. В мозге цитокины влияют на метаболизм нейромедиаторов, изменяя синтез и обратный захват серотонина, дофамина, норадреналина. Они модулируют активность гипоталамо-гипофизарно-надпочечниковой оси, влияя на продукцию кортизола. Они активируют микроглию — резидентные иммунные клетки мозга, — которая, в свою очередь, продуцирует локальные воспалительные медиаторы и влияет на функцию нейронов. Эти центральные эффекты цитокинов формируют характерный симптомокомплекс, известный как «болезненное поведение»: усталость, сонливость, снижение аппетита, социальная изоляция, ангедония, когнитивные нарушения. В эволюционном контексте болезненное поведение адаптивно: оно способствует экономии ресурсов и выздоровлению при острой инфекции. Однако при хроническом стрессе, когда воспаление персистирует без реальной инфекции, эти симптомы становятся проявлениями депрессии и хронической усталости.
Двунаправленность связи между мозгом и иммунной системой означает, что не только стресс влияет на иммунитет, но и состояние иммунной системы влияет на восприятие стресса и стресс-реактивность. Исследования демонстрируют, что экспериментальное повышение уровня провоспалительных цитокинов у здоровых добровольцев — путём введения низких доз бактериального эндотоксина или рекомбинантных цитокинов — вызывает симптомы депрессии, тревоги, когнитивные нарушения, даже в отсутствие субъективных признаков болезни. Хроническое воспаление, связанное с соматическими заболеваниями — ревматоидным артритом, воспалительными заболеваниями кишечника, ожирением, — ассоциировано с повышенным риском депрессии. Эти данные послужили основой для воспалительной гипотезы депрессии, предполагающей, что по крайней мере у части пациентов депрессия представляет собой проявление хронического воспаления и может отвечать на противовоспалительную терапию. Клинические испытания противовоспалительных препаратов при депрессии демонстрируют обнадёживающие, хотя и неоднородные результаты, особенно у пациентов с повышенными маркерами воспаления. Эта область активно развивается и имеет потенциал для персонализированного подхода к терапии, основанного на иммунном профиле пациента.
Микроглия — резидентные иммунные клетки центральной нервной системы — заслуживает особого упоминания как критически важное связующее звено между периферическим воспалением, стрессом и нейробиологическими изменениями. В отличие от большинства иммунных клеток, микроглия постоянно присутствует в мозге, составляя от пяти до десяти процентов всех клеток нервной ткани. В здоровом мозге микроглия выполняет «хозяйственные» функции: мониторирует нейронную активность, удаляет синаптический дебрис, участвует в ремоделировании синапсов. Однако при стрессе и воспалении микроглия активируется, переходя в провоспалительное состояние, меняя морфологию и продуцируя цитокины, хемокины, активные формы кислорода. Активированная микроглия может повреждать нейроны и синапсы, нарушать нейрогенез в гиппокампе, влиять на нейромедиаторный метаболизм. Исследования на животных моделях демонстрируют, что хронический стресс вызывает активацию микроглии в областях мозга, связанных с эмоциями и когнитивными функциями, и что блокада этой активации предотвращает некоторые поведенческие последствия стресса. Постмортемные исследования мозга людей с депрессией и суицидом выявляют признаки микроглиальной активации. Концепция нейровоспаления как механизма стресс-связанных нарушений мозга приобретает всё большее признание.
Иммунная система, таким образом, предстаёт не как изолированная защитная структура, но как интегральный компонент системы стресс-ответа, вносящий существенный вклад в адаптивные и патологические последствия стресса. Острый стресс мобилизует иммунную защиту в подготовке к возможной травме, что адаптивно в эволюционном контексте. Хронический стресс истощает адаптивный иммунитет, повышая восприимчивость к инфекциям, и одновременно активирует хроническое воспаление, повреждающее ткани и влияющее на мозг. Цитокины, продуцируемые периферическими иммунными клетками и активированной микроглией мозга, формируют психологические симптомы — усталость, ангедонию, когнитивные нарушения, — которые традиционно рассматривались как чисто «психические», но имеют отчётливый иммунобиологический компонент. Это понимание открывает новые терапевтические возможности: модуляция иммунной функции и воспаления может быть путём к облегчению психологических последствий стресса. Однако сложность иммунной системы и её тонкая настройка требуют осторожности: грубое подавление воспаления может ослабить необходимую защиту от патогенов. Персонализированный подход, учитывающий индивидуальный иммунный профиль и природу стресса, представляется наиболее перспективным направлением.
2.4 Митохондрии — энергетические станции под угрозой
Митохондрии занимают совершенно особое место среди клеточных органелл благодаря своей центральной роли в энергетическом метаболизме и, как стало ясно в последние десятилетия, благодаря множеству дополнительных функций, критически важных для жизни и смерти клетки. Эти небольшие органеллы, насчитывающие от нескольких сотен до нескольких тысяч на клетку в зависимости от её метаболических потребностей, являются основными производителями аденозинтрифосфата — универсальной энергетической валюты, необходимой практически для всех активных процессов в клетке. Однако представление о митохондриях исключительно как об «энергетических станциях» существенно устарело и не отражает всей полноты их биологического значения. Современные исследования демонстрируют, что митохондрии участвуют в регуляции внутриклеточной сигнализации, метаболизме кальция, продукции активных форм кислорода как сигнальных молекул, синтезе ряда ключевых метаболитов, и, что особенно важно в контексте стресса, в принятии «решения» о судьбе клетки — её выживании или программируемой гибели. Связь между митохондриями и стрессом носит глубокий и многогранный характер: с одной стороны, стресс предъявляет повышенные требования к энергопродукции и может повреждать митохондрии; с другой стороны, состояние митохондрий определяет способность клетки справляться со стрессом и во многом определяет его исход. Понимание митохондриальной биологии стресса открывает новые перспективы для объяснения механизмов стресс-связанных патологий и для разработки терапевтических стратегий.
Процесс производства энергии в митохондриях представляет собой замечательный пример молекулярной инженерии, в котором энергия, высвобождаемая при окислении питательных веществ, улавливается и преобразуется в химически удобную форму аденозинтрифосфата. Этот процесс, известный как окислительное фосфорилирование, протекает на внутренней митохондриальной мембране, которая образует многочисленные складки — кристы, — увеличивающие площадь поверхности для размещения белковых комплексов электрон-транспортной цепи. Электроны, извлечённые из молекул-переносчиков, образующихся при окислении глюкозы и жирных кислот в цикле трикарбоновых кислот, передаются по цепи белковых комплексов, и энергия этого переноса используется для выкачивания протонов через внутреннюю мембрану, создавая электрохимический градиент. Обратный поток протонов через фермент АТФ-синтазу приводит во вращение его каталитическую субъединицу и обеспечивает синтез аденозинтрифосфата из аденозиндифосфата и неорганического фосфата. Этот элегантный механизм чрезвычайно эффективен, однако не лишён побочных эффектов: часть электронов «утекает» из цепи и напрямую реагирует с кислородом, образуя супероксидный радикал и другие активные формы кислорода. В нормальных условиях эти побочные продукты нейтрализуются антиоксидантными системами, но при стрессе баланс может нарушаться.
Стресс предъявляет к митохондриям повышенные требования, которые могут превышать их адаптивные возможности и приводить к дисфункции. Активация стресс-ответа сопровождается резким увеличением энергетических потребностей организма: синтез стресс-индуцируемых белков, усиленная нейронная активность, мышечное напряжение, активация иммунных клеток — все эти процессы требуют дополнительного аденозинтрифосфата. Митохондрии отвечают на эти требования увеличением интенсивности окислительного фосфорилирования, что, однако, сопровождается повышенной продукцией активных форм кислорода. Одновременно гормоны стресса оказывают прямое влияние на митохондриальную функцию: глюкокортикоиды модулируют экспрессию генов, кодирующих компоненты дыхательной цепи, и влияют на динамику митохондрий; катехоламины через внутриклеточные сигнальные каскады также воздействуют на митохондриальный метаболизм. При остром стрессе эти изменения адаптивны и обратимы, но при хроническом стрессе кумулятивное воздействие повышенной нагрузки и окислительного повреждения может приводить к устойчивой митохондриальной дисфункции. Митохондрии начинают производить меньше энергии при большей «утечке» электронов, что создаёт порочный круг: энергодефицит ограничивает репаративные возможности клетки, а продолжающееся окислительное повреждение усугубляет дисфункцию.
Концепция митохондриальной аллостатической нагрузки, предложенная исследователями в последние годы, интегрирует понимание митохондриальной биологии с классической теорией аллостаза и аллостатической нагрузки. Подобно тому как аллостатическая нагрузка на уровне организма представляет собой кумулятивный износ физиологических систем вследствие хронической адаптации к стрессорам, митохондриальная аллостатическая нагрузка описывает накопление повреждений и дисфункции митохондрий в результате хронического энергетического стресса. Митохондрии особенно уязвимы к накоплению повреждений по ряду причин: они содержат собственную ДНК, расположенную в непосредственной близости от источника активных форм кислорода и не защищённую гистоновыми белками; их мембраны богаты ненасыщенными жирными кислотами, подверженными перекисному окислению; их белки постоянно экспонированы к окислительной среде. Мутации митохондриальной ДНК накапливаются с возрастом и при хроническом стрессе, приводя к синтезу дефектных компонентов дыхательной цепи и дальнейшему усугублению дисфункции. Эта концепция позволяет понять, как хронический стресс на клеточном уровне транслируется в ускоренное старение и повышенный риск возраст-ассоциированных заболеваний.
Нейроны представляют собой клетки с наивысшей зависимостью от митохондриальной функции и, соответственно, с наибольшей уязвимостью к митохондриальной дисфункции при стрессе. Мозг, составляя лишь около двух процентов массы тела, потребляет приблизительно двадцать процентов всего кислорода организма, и бо́льшая часть этой энергии расходуется нейронами на поддержание ионных градиентов, необходимых для электрической сигнализации, и на синаптическую передачу. Нейроны не способны переключаться на анаэробный метаболизм в той мере, в какой это могут делать другие клетки, и критически зависят от непрерывного снабжения кислородом и глюкозой. Более того, нейроны являются постмитотическими клетками — они не делятся во взрослом организме (за редкими исключениями), и, следовательно, накапливающиеся митохондриальные повреждения не могут быть «разбавлены» клеточным делением. Исследования демонстрируют, что хронический стресс вызывает митохондриальную дисфункцию в ключевых областях мозга — гиппокампе, префронтальной коре, миндалине — и что эта дисфункция коррелирует с когнитивными и эмоциональными нарушениями. Митохондриальная недостаточность может объяснять многие аспекты «мозгового тумана», усталости и когнитивного снижения, ассоциированных с хроническим стрессом.
Митохондриальная динамика — процессы слияния и деления митохондрий — представляет собой важный механизм адаптации к меняющимся энергетическим потребностям и контроля качества митохондриальной популяции. Митохондрии не являются статичными структурами, но постоянно сливаются друг с другом и делятся, формируя динамическую сеть, топология которой изменяется в зависимости от метаболического состояния клетки. Слияние позволяет обмениваться компонентами между митохондриями, «разбавляя» повреждения и компенсируя дефекты. Деление необходимо для размножения митохондрий при клеточном делении и для изоляции необратимо повреждённых митохондрий перед их удалением. Стресс оказывает существенное влияние на баланс между слиянием и делением: острый стресс часто вызывает усиление слияния, что может представлять защитную реакцию; хронический стресс, напротив, ассоциирован с избыточной фрагментацией митохондриальной сети, что снижает эффективность энергопродукции и способствует накоплению дисфункциональных органелл. Белки, регулирующие митохондриальную динамику, — митофузины, белок оптической атрофии первого типа, белок, связанный с динамином, — являются потенциальными мишенями для терапевтического воздействия при стресс-связанных нарушениях.
Митофагия — селективная аутофагия митохондрий — представляет собой критически важный механизм контроля качества, обеспечивающий удаление необратимо повреждённых митохондрий и предотвращающий их токсическое воздействие на клетку. Когда митохондрия утрачивает мембранный потенциал вследствие повреждения, на её поверхности накапливается киназа PINK1, которая в норме быстро деградирует в здоровых митохондриях. Накопление PINK1 рекрутирует убиквитинлигазу Паркин, которая убиквитинирует белки наружной мембраны митохондрии, маркируя её для поглощения аутофагосомой и последующей деградации в лизосоме. Мутации в генах PINK1 и Паркин ассоциированы с наследственными формами болезни Паркинсона, что подчёркивает критическую роль митофагии для здоровья нейронов. При хроническом стрессе эффективность митофагии может снижаться вследствие общего подавления аутофагии или специфических нарушений в компонентах митофагического пути. Это приводит к накоплению дисфункциональных митохондрий, которые продолжают генерировать активные формы кислорода, высвобождать проапоптотические факторы и потреблять клеточные ресурсы без адекватной продукции энергии. Стимуляция митофагии рассматривается как потенциальная терапевтическая стратегия при стресс-связанных и нейродегенеративных заболеваниях.
Активные формы кислорода, продуцируемые митохондриями, представляют собой двуликую реальность клеточной биологии — они могут быть как повреждающими агентами, так и важными сигнальными молекулами. Традиционный взгляд на активные формы кислорода исключительно как на токсичные побочные продукты метаболизма, подлежащие немедленной нейтрализации, уступил место более нюансированному пониманию. В умеренных концентрациях активные формы кислорода участвуют во внутриклеточной сигнализации, модулируя активность транскрипционных факторов, киназ, фосфатаз и других регуляторных белков через обратимое окисление критических цистеиновых остатков. Этот «редокс-сигналинг» вовлечён в регуляцию множества клеточных процессов, включая адаптацию к стрессу. Умеренный окислительный стресс активирует защитные программы — индукцию антиоксидантных ферментов, белков теплового шока, механизмов репарации — что может повышать устойчивость клетки к последующим воздействиям. Однако при избыточной продукции активных форм кислорода, превышающей буферную ёмкость антиоксидантных систем, начинается неконтролируемое окислительное повреждение белков, липидов и нуклеиновых кислот. Баланс между сигнальными и повреждающими эффектами активных форм кислорода зависит от их концентрации, локализации, продолжительности воздействия и состояния антиоксидантной защиты — параметров, которые существенно изменяются при хроническом стрессе.
Митохондрии играют центральную роль в регуляции программируемой клеточной гибели — апоптоза, — что связывает энергетический метаболизм с решениями о судьбе клетки при стрессе. Внутренний, или митохондриальный, путь апоптоза инициируется проапоптотическими членами семейства белков Bcl-2, которые при определённых сигналах олигомеризуются в наружной митохондриальной мембране и формируют поры, через которые из межмембранного пространства высвобождается цитохром С и другие апоптогенные факторы. Цитохром С в цитоплазме запускает каскад каспаз — протеаз, осуществляющих упорядоченную деградацию клетки. Антиапоптотические члены того же семейства Bcl-2 противодействуют этому процессу, и баланс между про- и антиапоптотическими сигналами определяет, погибнет клетка или выживет. Стресс модулирует этот баланс через множество механизмов: глюкокортикоиды регулируют экспрессию членов семейства Bcl-2; окислительный стресс может напрямую повреждать митохондриальные мембраны; дефицит энергии ослабляет репаративные процессы и склоняет баланс в сторону гибели. Понимание митохондриальной регуляции апоптоза критически важно для объяснения тканеспецифических последствий стресса — например, усиленной гибели нейронов гиппокампа при хроническом стрессе.
Терапевтические стратегии, направленные на поддержание митохондриальной функции при стрессе, привлекают растущее внимание исследователей и клиницистов. Антиоксиданты, таргетированные к митохондриям, — такие как соединения на основе трифенилфосфония, концентрирующиеся в митохондриях благодаря их положительному заряду и отрицательному мембранному потенциалу органелл, — демонстрируют защитные эффекты в экспериментальных моделях стресса. Коэнзим Q10, являющийся компонентом электрон-транспортной цепи, используется как добавка для поддержания митохондриальной функции, хотя клинические данные о его эффективности неоднородны. Стимуляция митохондриального биогенеза через активацию транскрипционного коактиватора PGC-1α рассматривается как стратегия увеличения митохондриального резерва; физическая активность является мощным естественным активатором этого пути. Кетогенная диета и интервальное голодание индуцируют метаболические адаптации, потенциально благоприятные для митохондриальной функции. Важно подчеркнуть, что большинство этих подходов находятся на стадии исследования, и их клиническая эффективность при стресс-связанных расстройствах требует дальнейшей верификации в контролируемых испытаниях. Тем не менее митохондрии представляются перспективной мишенью для вмешательств, направленных на повышение устойчивости к стрессу и смягчение его последствий.
2.5 Клеточное старение и программируемая гибель
Судьба клетки, подвергающейся стрессу, не ограничивается бинарным выбором между полным восстановлением и немедленной гибелью — между этими полюсами существует спектр промежуточных состояний, которые имеют критическое значение для понимания долгосрочных последствий стресса для организма. Клеточное старение, или сенесценция, представляет собой одно из таких состояний: клетка остаётся живой и метаболически активной, но необратимо утрачивает способность к делению и приобретает характерный секреторный фенотип, оказывающий влияние на окружающие ткани. Программируемая клеточная гибель, или апоптоз, представляет собой противоположный исход — упорядоченную самоликвидацию клетки, которая, однако, не всегда является патологическим событием, а во многих контекстах служит необходимым механизмом поддержания тканевого гомеостаза. Стресс влияет на оба эти процесса, сдвигая баланс между выживанием, старением и гибелью клеток и тем самым внося вклад в патогенез разнообразных заболеваний. Понимание клеточной сенесценции и апоптоза в контексте стресса позволяет по-новому взглянуть на механизмы ускоренного старения организма при хроническом стрессе и на возможности терапевтического вмешательства.
Клеточная сенесценция была первоначально описана как феномен репликативного старения — ограниченного числа делений, которое способна совершить нормальная клетка в культуре, — и связывалась с укорочением теломер при каждом клеточном делении. Теломеры — повторяющиеся нуклеотидные последовательности на концах хромосом — защищают хромосомы от деградации и слияния, но укорачиваются при репликации ДНК вследствие неспособности репликативного аппарата полностью копировать концевые участки. Когда теломеры достигают критически короткой длины, клетка воспринимает это как повреждение ДНК и активирует программу сенесценции, предотвращающую дальнейшие деления и потенциальную геномную нестабильность. Однако впоследствии было установлено, что сенесценция может индуцироваться множеством других стимулов независимо от теломерной длины: онкогенной активацией, окислительным стрессом, повреждением ДНК, митохондриальной дисфункцией, эпигенетическими нарушениями. Эти формы стресс-индуцированной сенесценции особенно релевантны для постмитотических или редко делящихся клеток и для понимания роли сенесценции в последствиях хронического стресса. Примечательно, что сам хронический психологический стресс ассоциирован с ускоренным укорочением теломер у людей — эффект, опосредованный, по-видимому, оксидативным стрессом и другими молекулярными механизмами.
Сенесцентные клетки характеризуются рядом морфологических и функциональных особенностей, которые позволяют их идентифицировать и которые определяют их влияние на окружающие ткани. Морфологически сенесцентные клетки часто увеличены в размерах, имеют уплощённую форму, характеризуются повышенной активностью ассоциированной со старением бета-галактозидазы — фермента, активность которого при определённых условиях выявления служит маркёром сенесценции. На молекулярном уровне сенесцентные клетки демонстрируют устойчивую активацию ингибиторов клеточного цикла p16INK4a и p21CIP1, которые блокируют прогрессию через контрольные точки клеточного цикла и обеспечивают необратимость ареста. Однако наиболее функционально значимой характеристикой сенесцентных клеток является их секреторный фенотип, ассоциированный со старением, — комплекс секретируемых факторов, включающий провоспалительные цитокины (интерлейкины-6 и -8, моноцитарный хемоаттрактантный белок-1), матриксные металлопротеиназы, факторы роста и другие биоактивные молекулы. Этот секреторный фенотип превращает сенесцентные клетки из пассивных «остановившихся» клеток в активных участников тканевой патологии, влияющих на окружающие клетки и способствующих хроническому воспалению.
Хронический стресс ускоряет накопление сенесцентных клеток в тканях через несколько взаимосвязанных механизмов, что вносит вклад в ускоренное биологическое старение и повышенный риск возраст-ассоциированных заболеваний. Окислительный стресс, сопровождающий хроническую активацию стресс-ответа, вызывает повреждения ДНК и митохондриальную дисфункцию, которые могут индуцировать сенесценцию независимо от репликативной истории клетки. Глюкокортикоиды оказывают сложное и контекстно-зависимое влияние на клеточную сенесценцию, которое может различаться в разных типах клеток и при разных режимах воздействия. Хроническое воспаление низкой интенсивности, характерное для хронического стресса, само по себе может индуцировать сенесценцию в соседних клетках через паракринное действие провоспалительных цитокинов — феномен, известный как «заражение» сенесценцией. Более того, сенесцентные клетки через свой секреторный фенотип поддерживают и усиливают воспаление, создавая порочный круг. Эпидемиологические исследования демонстрируют, что маркёры клеточной сенесценции, включая укорочение теломер и повышение экспрессии p16INK4a, ассоциированы с историей хронического стресса и психосоциального неблагополучия, подтверждая клиническую релевантность этих механизмов.
Метафора «клеток-зомби» получила широкое распространение для описания сенесцентных клеток и, несмотря на свою образность, довольно точно передаёт их биологическую сущность. Подобно мифическим зомби, сенесцентные клетки находятся в промежуточном состоянии между жизнью и смертью — они не выполняют нормальных функций живых клеток, но и не удаляются, как это происходит с погибшими клетками. Подобно зомби, которые, согласно легендам, «заражают» живых, сенесцентные клетки через свой секреторный фенотип могут индуцировать сенесценцию в соседних здоровых клетках, распространяя дисфункцию по ткани. Накопление «клеток-зомби» с возрастом и при хроническом стрессе рассматривается как один из фундаментальных механизмов старения организма. Эта концепция породила новое направление терапевтических исследований — разработку сенолитиков, препаратов, селективно уничтожающих сенесцентные клетки. Экспериментальные исследования на мышах демонстрируют, что удаление сенесцентных клеток с помощью генетических или фармакологических подходов увеличивает продолжительность здоровой жизни и уменьшает проявления возраст-ассоциированных заболеваний. Клинические испытания сенолитиков находятся на ранних стадиях, но представляют значительный интерес для потенциального применения при стресс-ассоциированном ускоренном старении.
Апоптоз, или программируемая клеточная гибель, представляет собой активный, энергозависимый процесс самоуничтожения клетки, который в норме является необходимым компонентом тканевого гомеостаза и развития. В отличие от некроза — неупорядоченной гибели клетки при грубом повреждении, сопровождающейся разрывом мембраны и воспалительной реакцией, — апоптоз протекает упорядоченно: клетка уменьшается в размерах, хроматин конденсируется, ДНК фрагментируется по межнуклеосомным линкерам, мембрана образует выпячивания, и в конечном счёте клетка распадается на апоптотические тельца, которые быстро поглощаются соседними клетками или макрофагами без развития воспаления. Этот процесс критически важен для элиминации повреждённых, инфицированных или потенциально опасных клеток, для формирования органов во время эмбрионального развития, для поддержания численности клеточных популяций в тканях. Нарушения апоптоза в обе стороны — как недостаточный, так и избыточный — ассоциированы с патологическими состояниями: недостаточный апоптоз способствует онкогенезу и аутоиммунным заболеваниям, избыточный — нейродегенерации и атрофии тканей.
Стресс оказывает сложное и тканеспецифичное влияние на апоптоз, которое зависит от типа клеток, характера и интенсивности стрессора, и временно́й динамики воздействия. В некоторых клеточных популяциях стресс усиливает апоптоз. Классическим примером является апоптоз тимоцитов — незрелых Т-лимфоцитов в тимусе — под действием глюкокортикоидов; этот эффект настолько выражен, что инволюция тимуса служит одним из признаков хронического стресса. В нейронах гиппокампа хронический стресс усиливает апоптоз, что вносит вклад в уменьшение объёма гиппокампа, наблюдаемое при депрессии и посттравматическом стрессовом расстройстве. В других клеточных популяциях стресс, напротив, может подавлять апоптоз: глюкокортикоиды оказывают антиапоптотическое действие на некоторые типы клеток, включая определённые популяции нейронов. Эта тканевая специфичность определяется различиями в экспрессии изоформ глюкокортикоидного рецептора, в балансе про- и антиапоптотических членов семейства Bcl-2, в активности сигнальных путей, регулирующих выживание клеток. Понимание этих различий необходимо для предсказания последствий стресса для конкретных органов и систем.
Нарушение баланса между апоптозом, сенесценцией и выживанием клеток при хроническом стрессе имеет далеко идущие последствия для тканевого гомеостаза и функции органов. В тканях с высоким пролиферативным потенциалом, таких как эпителий кишечника или костный мозг, усиленный апоптоз может нарушать обновление клеточных популяций и вести к атрофии или дисфункции. В тканях с ограниченной регенеративной способностью, таких как нервная ткань или миокард, потеря клеток через апоптоз является необратимой и приводит к прогрессирующему снижению функционального резерва. Одновременно накопление сенесцентных клеток в тканях создаёт провоспалительную среду, нарушающую функцию оставшихся здоровых клеток. Хронический стресс может приводить к парадоксальной ситуации, когда повреждённые клетки, которые должны были бы погибнуть, переходят в сенесценцию и персистируют, тогда как здоровые клетки подвергаются избыточному апоптозу вследствие паракринных эффектов сенесцентных соседей. Распутывание этих сложных взаимоотношений и понимание факторов, определяющих судьбу конкретной клетки при стрессе, представляет собой активную область исследований.
Терапевтические импликации понимания клеточной сенесценции и апоптоза при стрессе включают несколько направлений. Сенолитики — препараты, селективно индуцирующие апоптоз в сенесцентных клетках, которые устойчивы к обычным проапоптотическим сигналам благодаря повышенной экспрессии антиапоптотических белков, — находятся на стадии активной разработки и ранних клинических испытаний. Сеноморфики — препараты, подавляющие секреторный фенотип сенесцентных клеток без их уничтожения, — представляют альтернативный подход. Антиоксиданты и другие агенты, предотвращающие индукцию сенесценции, могут рассматриваться как превентивная стратегия. С другой стороны, вмешательства, модулирующие апоптоз, должны учитывать тканевую специфичность и избегать как усиления апоптоза в уязвимых популяциях, так и его подавления в клетках, которые должны быть элиминированы. Интегративный подход, сочетающий снижение стрессовой нагрузки с поддержанием клеточных защитных механизмов и, возможно, селективным удалением накопившихся сенесцентных клеток, представляется наиболее перспективным направлением для противодействия клеточным последствиям хронического стресса.
2.6 Межклеточная коммуникация — клетки как социальная сеть
Представление об организме как о совокупности индивидуальных клеток, каждая из которых автономно выполняет свои функции, является глубоким упрощением, не отражающим реальной сложности биологических систем. В действительности клетки многоклеточного организма непрерывно взаимодействуют друг с другом через множество каналов коммуникации, формируя сложную сеть, свойства которой несводимы к свойствам отдельных элементов. Эта межклеточная коммуникация обеспечивает координацию функций триллионов клеток, их согласованный ответ на внешние и внутренние сигналы, поддержание тканевой архитектуры и гомеостаза. В контексте стресса межклеточная коммуникация приобретает особое значение: она обеспечивает распространение сигнала тревоги от клеток, непосредственно воспринимающих стрессор, к отдалённым клеткам и тканям; она координирует системный ответ организма; она модулирует индивидуальные клеточные реакции в зависимости от сигналов от соседей и от отдалённых регуляторных центров. Однако та же межклеточная коммуникация может становиться механизмом патологии, когда аберрантные сигналы от повреждённых или дисфункциональных клеток распространяются по организму, создавая хроническое стрессовое воздействие на здоровые ткани. Метафора клеток как участников социальной сети продуктивна для понимания этих процессов: подобно тому как информация распространяется в социальных сетях, влияя на настроение и поведение людей, молекулярные сигналы распространяются между клетками, влияя на их функцию и судьбу.
Классические механизмы межклеточной коммуникации включают несколько модальностей, различающихся по расстоянию действия, скорости и специфичности передаваемых сигналов. Паракринная сигнализация предполагает секрецию сигнальных молекул, которые диффундируют через внеклеточное пространство и воздействуют на соседние клетки, находящиеся в непосредственной близости от источника сигнала. Этот механизм опосредует локальную координацию клеточных ответов и играет важную роль в воспалительных процессах, заживлении ран, развитии тканей. Эндокринная сигнализация, подробно рассмотренная ранее, предполагает выделение гормонов в кровоток и их действие на отдалённые клетки-мишени. Юкстакринная сигнализация осуществляется при непосредственном контакте между клетками через мембранные белки, которые взаимодействуют с рецепторами на поверхности соседней клетки. Синаптическая передача, специфичная для нервной системы, обеспечивает высокоточную и быструю коммуникацию между нейронами через специализированные контакты — синапсы. Каждый из этих механизмов задействуется при стрессе, и их интеграция обеспечивает координированный ответ организма на угрозу.
Щелевые контакты, или «gap junctions», представляют собой уникальную форму межклеточной коммуникации, обеспечивающую прямую цитоплазматическую связь между соседними клетками. Эти структуры образованы белками коннексинами, которые формируют каналы, пронизывающие мембраны обеих контактирующих клеток и создающие непрерывный проход между их цитоплазмами. Через щелевые контакты могут проходить малые молекулы с массой до примерно одного килодальтона, включая ионы, вторичные посредники (циклический аденозинмонофосфат, инозитолтрифосфат, кальций), метаболиты, нуклеотиды. Это позволяет клеткам обмениваться сигнальной и метаболической информацией практически мгновенно, синхронизируя свои ответы. В сердце щелевые контакты обеспечивают электрическую связь между кардиомиоцитами, необходимую для синхронного сокращения. В нервной системе щелевые контакты между определёнными популяциями нейронов и между нейронами и глиальными клетками участвуют в синхронизации активности и в метаболической поддержке нейронов. Стресс влияет на экспрессию и функцию коннексинов, что может нарушать межклеточную координацию; изменения щелевых контактов наблюдаются при сердечных аритмиях, ассоциированных со стрессом, и при нейропсихиатрических расстройствах.
Экзосомы и другие внеклеточные везикулы представляют собой относительно недавно охарактеризованный и чрезвычайно важный механизм межклеточной коммуникации, который радикально изменил понимание того, как клетки обмениваются информацией. Экзосомы — это мембранные пузырьки диаметром от тридцати до ста пятидесяти нанометров, которые секретируются практически всеми типами клеток и обнаруживаются во всех биологических жидкостях организма. Они образуются путём отпочковывания от внутриклеточных мембран и последующего высвобождения при слиянии мультивезикулярных телец с плазматической мембраной. Ключевая особенность экзосом состоит в том, что они несут не просто случайную выборку клеточного содержимого, но специфически отобранный «груз»: определённые белки, липиды, и, что особенно важно, нуклеиновые кислоты, включая информационные РНК и микроРНК. Микро-РНК, доставленные экзосомами в клетку-мишень, могут регулировать экспрессию генов в этой клетке, что представляет собой форму горизонтального переноса регуляторной информации между клетками. Экзосомы способны путешествовать на большие расстояния через кровоток и другие биологические жидкости, обеспечивая коммуникацию между отдалёнными тканями.
Стресс существенно изменяет экзосомную коммуникацию, модулируя как продукцию везикул, так и их молекулярный состав. Исследования демонстрируют, что клетки, подвергающиеся стрессу, выделяют больше экзосом, и эти экзосомы несут характерный «стрессовый» профиль содержимого. Они могут содержать белки теплового шока, которые, будучи высвобождены внеклеточно, могут активировать иммунные клетки и способствовать воспалительному ответу. Они несут изменённый набор микро-РНК, которые могут модулировать стресс-ответ в клетках-мишенях. Циркулирующие экзосомы могут служить системой дистанционного оповещения, информирующей отдалённые ткани о стрессе в определённом органе. Однако эта система может также распространять патологические сигналы: экзосомы от воспалённых или повреждённых клеток могут индуцировать воспаление и дисфункцию в здоровых тканях. Экзосомы от сенесцентных клеток несут компоненты их секреторного фенотипа и могут индуцировать сенесценцию в соседних клетках. Таким образом, экзосомная коммуникация представляет собой двусторонний меч, способный как координировать адаптивный ответ, так и распространять патологию.
Иммунные клетки играют особую роль в межклеточной коммуникации при стрессе благодаря своей мобильности и способности продуцировать широкий спектр сигнальных молекул. В отличие от большинства клеток, которые фиксированы в своих тканях, иммунные клетки активно мигрируют по организму, получая информацию в одних тканях и доставляя её в другие. Макрофаги, инфильтрирующие ткань, получают сигналы от локальных клеток о повреждении или стрессе и в ответ продуцируют цитокины, которые влияют на функцию соседних клеток и привлекают дополнительные иммунные клетки. Активированные макрофаги и другие иммунные клетки могут мигрировать в лимфатические узлы, где они взаимодействуют с лимфоцитами и координируют системный иммунный ответ. Цитокины, продуцируемые иммунными клетками, поступают в системную циркуляцию и влияют на отдалённые органы, включая мозг. Эта особая роль иммунных клеток как «путешествующих информаторов» объясняет, почему иммунная система является ключевым связующим звеном между локальными повреждениями и системным стресс-ответом, и почему хроническое воспаление имеет столь широкие системные последствия.
Внеклеточный матрикс, традиционно рассматривавшийся как пассивный каркас для клеток, в действительности представляет собой активного участника межклеточной коммуникации. Матрикс состоит из сложной сети белков — коллагенов, эластина, фибронектина, ламинина — и полисахаридов, которые секретируются клетками и формируют трёхмерную среду, в которой клетки живут. Матрикс не только обеспечивает механическую поддержку, но и содержит связанные факторы роста и другие сигнальные молекулы, которые могут высвобождаться при ремоделировании матрикса. Физические свойства матрикса — его жёсткость, порозность, топография — воспринимаются клетками через механосенсорные рецепторы, называемые интегринами, и влияют на клеточное поведение, включая пролиферацию, миграцию и дифференцировку. Стресс вызывает ремоделирование матрикса: провоспалительные цитокины индуцируют продукцию матриксных металлопротеиназ, расщепляющих компоненты матрикса; фиброгенные факторы стимулируют избыточное отложение коллагена. Эти изменения матрикса модулируют сигнальную среду для всех клеток ткани и могут способствовать персистенции патологических состояний даже после устранения первоначального стрессора.
Нарушения межклеточной коммуникации при хроническом стрессе могут создавать порочные круги, в которых аберрантные сигналы самоподдерживаются и усиливаются. Классическим примером является воспалительный порочный круг: повреждённые клетки продуцируют провоспалительные цитокины; эти цитокины активируют соседние клетки и привлекают иммунные клетки; активированные клетки продуцируют ещё больше цитокинов; хроническое воспаление повреждает дополнительные клетки, замыкая круг. Сенесцентные клетки через свой секреторный фенотип индуцируют сенесценцию в соседях, распространяя дисфункцию по ткани. Нарушенная коммуникация между нейронами и глией в мозге при хроническом стрессе может приводить к недостаточной метаболической и трофической поддержке нейронов и усугублять нейрональную дисфункцию. Экзосомы от стрессированных клеток переносят патологические сигналы к здоровым клеткам, вовлекая их в дисфункциональное состояние. Разрыв этих порочных кругов представляет собой важную терапевтическую задачу, решение которой требует понимания сетевой организации межклеточной коммуникации и идентификации ключевых узлов, воздействие на которые может нормализовать системную динамику.
Понимание организма как сети взаимодействующих клеток имеет глубокие импликации для концептуализации стресса и для подходов к его терапии. На индивидуальном уровне это означает, что стресс, первоначально локализованный в определённых клетках или тканях, неизбежно распространяется через сигнальные сети и влияет на весь организм. Хронический психологический стресс, воспринимаемый мозгом, через нейроэндокринные и нейроиммунные пути достигает периферических тканей и вызывает молекулярные изменения на клеточном уровне. Локальное повреждение или инфекция через продукцию цитокинов и циркулирующих экзосом влияет на отдалённые органы и на мозг, модулируя поведение и эмоциональное состояние. На терапевтическом уровне это означает, что эффективные вмешательства могут таргетировать межклеточную коммуникацию: блокада провоспалительных цитокинов может прервать патологические сигнальные каскады; модуляция экзосомной секреции или захвата может влиять на распространение стрессовых сигналов; восстановление нормального состава внеклеточного матрикса может нормализовать сигнальную среду для резидентных клеток. Сетевая перспектива также подчёркивает значимость превентивных вмешательств: предотвращение инициации патологических коммуникационных каскадов более эффективно, чем попытки прервать уже установившиеся порочные круги.
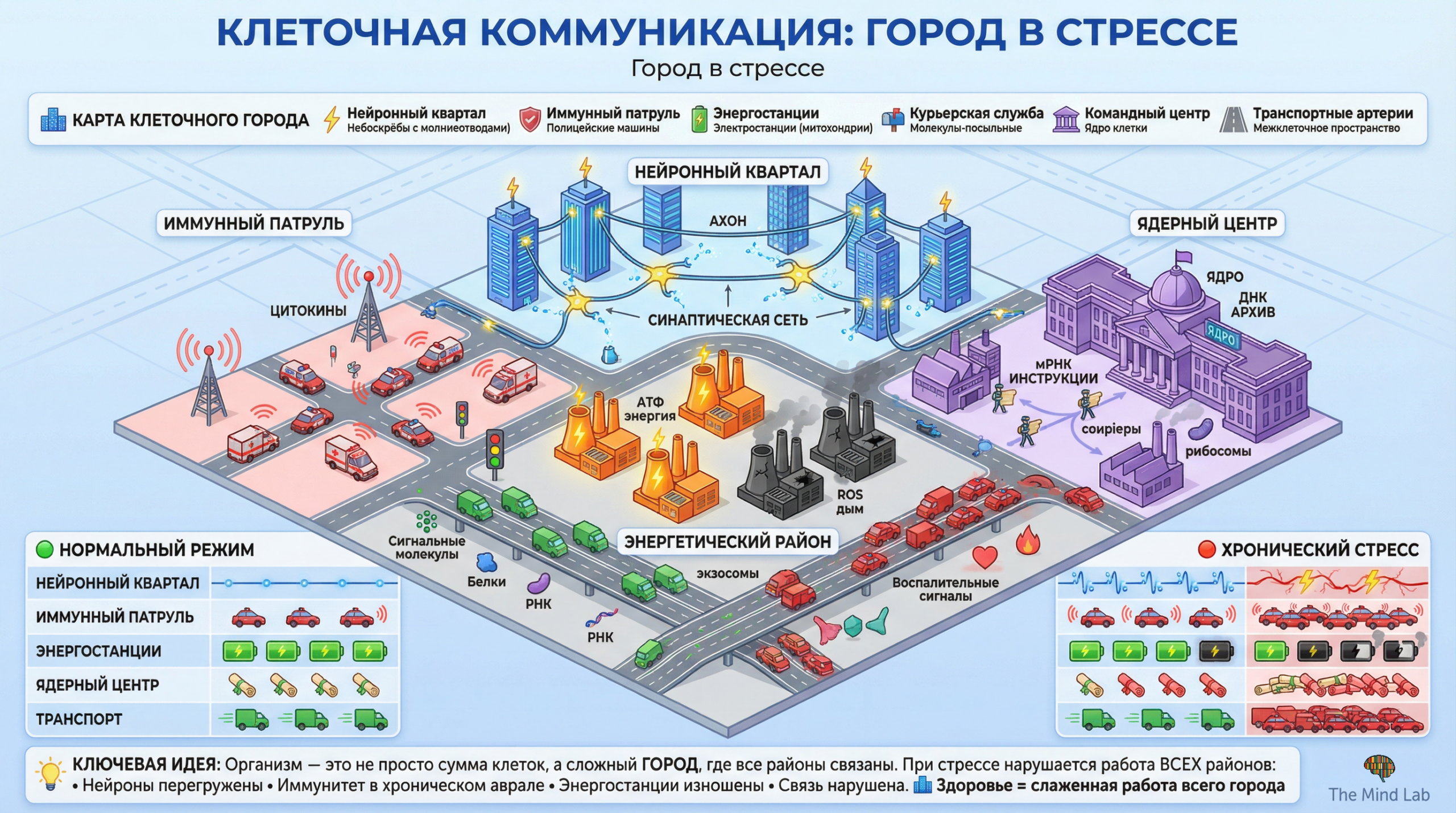
3. Системный уровень — интеграция и регуляция
3.1 Три главные системы-регулятора стресса
Переход от клеточного уровня анализа к системному представляет собой качественный скачок в понимании стресса, поскольку на этом уровне мы имеем дело не просто с совокупностью индивидуальных клеточных реакций, но с координированной деятельностью специализированных физиологических систем, обеспечивающих интегрированный ответ целого организма. Три системы — нервная, эндокринная и иммунная — традиционно выделяются как главные регуляторы стресс-ответа, каждая из которых вносит свой уникальный вклад в адаптацию организма к угрозам и вызовам. Однако современное понимание физиологии стресса решительно отходит от представления об этих системах как об автономных сущностях, работающих параллельно и независимо друг от друга. Исследования последних десятилетий убедительно продемонстрировали, что нервная, эндокринная и иммунная системы образуют единую интегрированную сеть, в которой каждый компонент непрерывно влияет на другие через множество каналов коммуникации. Эта концепция нейроэндокриноиммунной интеграции фундаментально изменила подход к изучению стресса и породила новые междисциплинарные области, такие как психонейроэндокринология и психонейроиммунология. Понимание этого интегративного принципа составляет необходимую основу для осмысления всех последующих аспектов науки о стрессе.
Нервная система обеспечивает самый быстрый канал коммуникации в организме, позволяя передавать сигналы от мозга к периферическим органам и обратно в течение миллисекунд. Эта скорость достигается благодаря электрохимической природе нервной передачи: электрические импульсы распространяются вдоль нервных волокон, а в местах контактов между нейронами и между нейронами и эффекторными клетками преобразуются в химические сигналы — нейромедиаторы. Нервная система первой реагирует на восприятие стрессора, мгновенно изменяя работу сердца, сосудов, дыхательной системы, желудочно-кишечного тракта и других органов. Симпатический отдел вегетативной нервной системы активирует реакцию «бей или беги», подготавливая организм к интенсивному физическому действию, тогда как парасимпатический отдел обеспечивает восстановление после стресса и поддержание базовых функций в состоянии покоя. Помимо прямых эффектов на органы, нервная система запускает каскады в двух других регуляторных системах: через гипоталамус она активирует эндокринный ответ, а через иннервацию лимфоидных органов и прямое выделение нейромедиаторов вблизи иммунных клеток — модулирует иммунную функцию. Таким образом, нервная система выступает не только как самостоятельный эффектор, но и как инициатор и координатор системного стресс-ответа.
Эндокринная система, в отличие от нервной, использует химические сигналы — гормоны, — которые секретируются железами внутренней секреции в кровоток и достигают клеток-мишеней по всему организму. Этот способ коммуникации медленнее нервной передачи — от минут до часов вместо миллисекунд, — однако он обеспечивает более устойчивые и системные эффекты, затрагивающие одновременно множество органов и тканей. Центральным элементом эндокринного стресс-ответа является гипоталамо-гипофизарно-надпочечниковая ось, которая обеспечивает продукцию глюкокортикоидов — гормонов с широчайшим спектром метаболических, иммунных и поведенческих эффектов. Однако стресс затрагивает и другие эндокринные оси: гипоталамо-гипофизарно-тиреоидную, регулирующую метаболизм; гипоталамо-гипофизарно-гонадную, регулирующую репродукцию; ось гормона роста; пролактиновую ось. Эндокринная система находится в постоянном диалоге с нервной: гипоталамус, являющийся частью мозга, переводит нервные сигналы в эндокринные команды; гормоны, в свою очередь, проникают в мозг и модулируют нейронную активность. С иммунной системой эндокринная связана не менее тесно: глюкокортикоиды являются мощными модуляторами иммунной функции, а цитокины, продуцируемые иммунными клетками, способны активировать гипоталамо-гипофизарно-надпочечниковую ось.
Иммунная система завершает триаду главных регуляторов стресса, и её включение в этот список отражает революционное изменение в понимании её функций, произошедшее за последние полвека. Традиционно иммунная система рассматривалась исключительно как механизм защиты от патогенов — бактерий, вирусов, паразитов — и от опухолевых клеток. Однако стало ясно, что иммунные клетки выполняют также важнейшие регуляторные функции, влияющие на работу нервной системы, метаболизм, поведение и психологическое состояние. Цитокины — сигнальные белки, продуцируемые иммунными клетками — способны достигать мозга и вызывать характерные изменения в нейромедиаторном метаболизме, приводящие к усталости, снижению мотивации, социальной изоляции, нарушениям сна и аппетита. Эти эффекты, объединяемые термином «болезненное поведение», изначально адаптивны — они способствуют экономии ресурсов для борьбы с инфекцией, — но при хроническом стрессе превращаются в источник страдания. Иммунная система экспрессирует рецепторы к гормонам стресса и нейромедиаторам, что делает её чувствительной к сигналам от двух других регуляторных систем. Взаимосвязь между мозгом и иммунитетом оказалась настолько глубокой, что породила целую дисциплину — психонейроиммунологию, — изучающую эту интеграцию.
Метафора оркестра продуктивна для понимания того, как три регуляторные системы координируют свою деятельность при стресс-ответе, и для осмысления последствий нарушения этой координации. Подобно тому как в симфоническом оркестре струнные, духовые и ударные инструменты играют различные партии, которые объединяются в единое музыкальное произведение под управлением дирижёра, нервная, эндокринная и иммунная системы реализуют различные аспекты стресс-ответа, интегрируемые мозгом в целостную адаптивную реакцию. Каждая секция оркестра имеет свой тембр и свои возможности: струнные способны к быстрым пассажам и тонким нюансам динамики, подобно нервной системе с её скоростью и точностью; духовые обеспечивают мощное и продолжительное звучание, подобно эндокринной системе с её устойчивыми системными эффектами; ударные задают ритм и создают акценты, подобно иммунной системе с её способностью усиливать и модулировать ответы других систем. Когда оркестр слаженно исполняет произведение под руководством опытного дирижёра, результатом является гармоничное звучание; когда координация нарушается, возникает какофония. Аналогично, адаптивный стресс-ответ требует согласованной работы всех трёх систем; дисрегуляция любой из них или нарушение их интеграции приводит к патологическим последствиям.
Взаимодействие между тремя регуляторными системами осуществляется через множество молекулярных каналов, которые образуют сложную сеть с многочисленными точками пересечения и петлями обратной связи. Нервная система влияет на иммунную через прямую иннервацию лимфоидных органов — тимуса, селезёнки, лимфатических узлов, костного мозга — симпатическими волокнами, высвобождающими норадреналин в непосредственной близости от иммунных клеток. Блуждающий нерв, основной компонент парасимпатической системы, реализует так называемый воспалительный рефлекс, подавляя избыточное воспаление через высвобождение ацетилхолина. Эндокринная система влияет на иммунную через глюкокортикоиды, которые связываются с рецепторами практически всех типов иммунных клеток и модулируют их функцию, а также через катехоламины надпочечников, действующие через адренорецепторы на иммунных клетках. Иммунная система влияет на нервную и эндокринную через цитокины, которые достигают мозга несколькими путями: через области с проницаемым гематоэнцефалическим барьером, через активный транспорт, через активацию афферентных нервов (особенно блуждающего), через индукцию вторичных медиаторов в эндотелии мозговых сосудов. Эти многообразные пути коммуникации обеспечивают непрерывный обмен информацией между системами.
Историческое развитие понимания нейроэндокриноиммунной интеграции прошло путь от изолированного изучения каждой системы к признанию их фундаментальной взаимосвязи. На протяжении большей части двадцатого века иммунология, эндокринология и нейронауки развивались как относительно независимые дисциплины со своими методами, концептуальными рамками и профессиональными сообществами. Представление об автономности иммунной системы было особенно укоренившимся: считалось, что иммунные реакции разворачиваются по собственным законам, недоступным для влияния со стороны мозга и психики. Работы Ганса Селье в середине века продемонстрировали связь между стрессом и иммунной функцией — в частности, инволюцию тимуса при хроническом стрессе, — однако механизмы этой связи оставались неясными. Прорыв произошёл в семидесятых-восьмидесятых годах благодаря нескольким ключевым открытиям: обнаружению иннервации лимфоидных органов, идентификации рецепторов к гормонам и нейромедиаторам на иммунных клетках, демонстрации влияния цитокинов на мозг, экспериментальным доказательствам условнорефлекторной модуляции иммунитета. Эти открытия заложили основу для формирования психонейроиммунологии как признанной научной дисциплины и для современного интегративного понимания стресса.
Концепция единой нейроэндокриноиммунной сети имеет принципиальное значение для понимания как адаптивных, так и патологических аспектов стресса. В адаптивном контексте интеграция систем обеспечивает координированный ответ, в котором каждый компонент дополняет и усиливает другие. Нервная система мгновенно мобилизует организм для немедленного действия; эндокринная обеспечивает устойчивую метаболическую поддержку этой мобилизации; иммунная готовится к возможным повреждениям и инфекциям. После устранения угрозы те же механизмы координации обеспечивают согласованное возвращение к исходному состоянию. В патологическом контексте нарушение интеграции между системами или дисфункция любого из компонентов сети приводит к несогласованности ответа. Хроническая активация симпатической нервной системы при ослабленной парасимпатической регуляции; нарушение обратной связи в гипоталамо-гипофизарно-надпочечниковой оси с хроническим повышением или, напротив, притуплением продукции кортизола; хроническое воспаление низкой интенсивности, поддерживаемое иммунными механизмами и влияющее на мозг — все эти паттерны отражают дезинтеграцию нейроэндокриноиммунной сети и лежат в основе различных стресс-ассоциированных расстройств.
Практические следствия понимания нейроэндокриноиммунной интеграции затрагивают как диагностику, так и терапию стресс-связанных состояний. В диагностическом отношении это означает, что оценка стресс-статуса пациента не может ограничиваться измерением одного параметра — например, уровня кортизола, — но требует комплексной оценки маркёров всех трёх систем и понимания их взаимоотношений. Высокий кортизол в сочетании с повышенными маркёрами воспаления может указывать на нарушение противовоспалительного действия глюкокортикоидов — так называемую глюкокортикоидную резистентность, — требующее иной терапевтической стратегии, чем высокий кортизол с нормальными воспалительными маркёрами. В терапевтическом отношении понимание интеграции открывает множество потенциальных точек вмешательства: воздействие на вегетативную нервную систему через дыхательные техники или стимуляцию блуждающего нерва может снижать воспаление; противовоспалительная терапия может улучшать психологическое состояние; психотерапия, изменяющая когнитивную оценку стрессоров, влияет на эндокринные и иммунные параметры. Интегративный подход к терапии стресса, учитывающий все три системы и их взаимодействия, представляется наиболее перспективным.
3.2 Нервная система — молниеносная передача команд
Нервная система представляет собой наиболее быстродействующую из трёх регуляторных систем стресс-ответа, обеспечивая передачу сигналов от органов чувств к мозгу и от мозга к эффекторным органам в течение миллисекунд. Эта скорость критически важна для выживания: когда хищник атакует или машина неожиданно выезжает на пешеходный переход, организм не может позволить себе ждать минуты или часы, пока гормоны достигнут своих мишеней — реакция должна быть мгновенной. Нервная система обеспечивает именно такую мгновенную реактивность благодаря электрической природе проведения сигнала вдоль нервных волокон и высокоспециализированной химической передаче в синапсах. Анатомически нервная система подразделяется на центральную, включающую головной и спинной мозг, и периферическую, включающую нервы, связывающие центральную нервную систему со всеми органами и тканями тела. Функционально периферическая нервная система делится на соматическую, управляющую произвольными движениями и обеспечивающую сознательное восприятие, и вегетативную, или автономную, регулирующую функции внутренних органов преимущественно без участия сознания. Именно вегетативная нервная система играет центральную роль в немедленном стресс-ответе, мгновенно изменяя работу сердца, сосудов, дыхательной системы и множества других органов.
Вегетативная нервная система исторически рассматривалась как «автономная» именно потому, что её функции осуществляются независимо от сознательного контроля и волевых усилий: мы не можем по желанию замедлить сердцебиение или остановить перистальтику кишечника так, как можем поднять руку или повернуть голову. Однако термин «автономная» не означает независимости от центральной нервной системы — напротив, вегетативные функции находятся под постоянным контролем высших отделов мозга, включая гипоталамус, лимбические структуры и даже кору больших полушарий. Вегетативная нервная система подразделяется на два главных отдела — симпатический и парасимпатический, — которые, как правило, оказывают противоположные влияния на органы-мишени и находятся в состоянии динамического баланса. Симпатический отдел активирует организм, подготавливая его к интенсивной деятельности: он ускоряет сердечный ритм, повышает артериальное давление, расширяет бронхи, перераспределяет кровоток от пищеварительной системы к мышцам, расширяет зрачки, стимулирует потоотделение. Парасимпатический отдел, напротив, способствует восстановлению и накоплению ресурсов: замедляет сердечный ритм, стимулирует пищеварение и секрецию пищеварительных соков, сужает бронхи и зрачки, способствует опорожнению мочевого пузыря и кишечника.
Классическая концепция реакции «бей или беги», сформулированная американским физиологом Уолтером Кэнноном в начале двадцатого века, описывает немедленную активацию симпатической нервной системы в ответ на восприятие угрозы. Кэннон наблюдал, как животные, столкнувшиеся с опасностью, демонстрируют комплекс физиологических изменений: учащение пульса, повышение давления, расширение зрачков, прекращение пищеварения, выброс глюкозы в кровь. Все эти изменения функционально направлены на обеспечение максимальной готовности к интенсивному физическому действию — либо борьбе с угрозой, либо бегству от неё. Симпатическая активация при стрессе не ограничивается влиянием на отдельные органы, но носит генерализованный характер благодаря анатомическим и нейрохимическим особенностям этой системы. Преганглионарные нейроны симпатической системы расположены в грудном и поясничном отделах спинного мозга и образуют синапсы с постганглионарными нейронами в паравертебральных ганглиях, которые связаны между собой в единую цепь. Это обеспечивает возможность широкой иррадиации возбуждения. Кроме того, симпатическая активация стимулирует мозговое вещество надпочечников к выбросу адреналина в кровь, что усиливает и пролонгирует симпатические эффекты через гуморальный механизм.
Парасимпатический отдел вегетативной нервной системы организован принципиально иначе и обеспечивает более дискретные, органоспецифичные влияния. Преганглионарные парасимпатические нейроны расположены в стволе мозга и в крестцовом отделе спинного мозга, а постганглионарные нейроны находятся непосредственно в стенках органов-мишеней или вблизи них. Блуждающий нерв, или вагус, является главным проводником парасимпатических влияний на большинство внутренних органов — сердце, лёгкие, желудочно-кишечный тракт вплоть до поперечной ободочной кишки, печень, поджелудочную железу. Стивен Порджес, современный исследователь вегетативной регуляции, предложил поливагальную теорию, выделяющую в составе блуждающего нерва эволюционно более древнюю немиелинизированную ветвь, связанную с реакциями замирания и выключения при непреодолимой угрозе, и более новую миелинизированную ветвь, связанную с социальной коммуникацией и состояниями спокойствия и контакта. Согласно этой теории, три уровня вегетативной регуляции — социальное взаимодействие, мобилизация (бей/беги) и иммобилизация (замирание) — последовательно активируются в зависимости от воспринимаемого уровня безопасности или угрозы, и терапия стресс-связанных расстройств должна учитывать этот иерархический принцип.
Метафора «педали газа» для симпатической системы и «педали тормоза» для парасимпатической интуитивно понятна, однако представляет некоторое упрощение реальных взаимоотношений между этими отделами. В действительности симпатическая и парасимпатическая системы не просто противодействуют друг другу, но находятся в сложных реципрокных и синергических взаимоотношениях в зависимости от органа и функционального контекста. В сердце два отдела действительно антагонистичны: симпатическая активация ускоряет ритм, парасимпатическая — замедляет. Однако для некоторых органов характерно преимущественное влияние одного отдела с минимальным вкладом другого. В ряде ситуаций наблюдается коактивация обоих отделов, а не реципрокные отношения. Понятие вегетативного баланса, или симпато-вагального баланса, отражает относительный вклад двух отделов в регуляцию и является важным интегративным показателем состояния организма. В покое у здорового человека преобладает парасимпатический тонус, обеспечивающий восстановительные процессы. При стрессе баланс сдвигается в сторону симпатической активации. Хронический стресс характеризуется устойчивым сдвигом баланса в симпатическую сторону — постоянно «нажатой педалью газа» — что истощает адаптивные резервы и повышает риск сердечно-сосудистых заболеваний.
Вариабельность сердечного ритма представляет собой неинвазивный метод оценки вегетативного баланса, приобретающий всё большее значение в исследованиях стресса и в клинической практике. Здоровое сердце не бьётся с метрономической регулярностью — интервалы между последовательными сокращениями постоянно варьируют под влиянием вегетативных модуляций. Парасимпатическая активация, опосредованная блуждающим нервом, вызывает быстрые колебания сердечного ритма, синхронизированные с дыханием (так называемая дыхательная синусовая аритмия: ритм ускоряется на вдохе и замедляется на выдохе). Симпатическая активация вызывает более медленные колебания. Анализ спектра колебаний сердечного ритма позволяет количественно оценить вклад симпатического и парасимпатического отделов. Высокая вариабельность сердечного ритма, особенно её высокочастотный компонент, отражающий вагальную активность, ассоциирована с хорошим здоровьем, психологическим благополучием, способностью к эмоциональной регуляции. Низкая вариабельность, напротив, является предиктором сердечно-сосудистых заболеваний, депрессии, тревожных расстройств, посттравматического стрессового расстройства. Тренировка вариабельности сердечного ритма с использованием биологической обратной связи рассматривается как перспективный метод улучшения вегетативной регуляции и повышения устойчивости к стрессу.
Центральная регуляция вегетативных функций осуществляется иерархической системой структур, включающей спинной мозг, ствол мозга, гипоталамус, лимбические структуры и префронтальную кору. Спинальный уровень обеспечивает базовые рефлекторные дуги, замыкающиеся внутри сегмента. Стволовые центры — ядра блуждающего нерва, ядра солитарного тракта, вентролатеральный отдел продолговатого мозга — интегрируют информацию от внутренних органов и координируют висцеральные рефлексы, включая барорефлекс, регулирующий артериальное давление. Гипоталамус представляет собой ключевой интегратор, связывающий вегетативную регуляцию с эндокринной, терморегуляторной, пищевой, питьевой и репродуктивной функциями; электрическая стимуляция различных областей гипоталамуса вызывает паттерны вегетативных реакций, соответствующих различным эмоциональным и мотивационным состояниям. Лимбические структуры — миндалина, гиппокамп, поясная кора — придают вегетативным реакциям эмоциональную окраску и контекстуальную зависимость; именно миндалина обеспечивает мгновенную вегетативную активацию при восприятии угрозы. Префронтальная кора осуществляет нисходящий контроль над лимбическими структурами и может модулировать вегетативные реакции на основе когнитивной оценки ситуации; нарушение этого контроля при хроническом стрессе ведёт к избыточным и неадекватным вегетативным ответам.
Последствия хронической вегетативной дисрегуляции при длительном стрессе затрагивают практически все системы организма и вносят существенный вклад в патогенез стресс-ассоциированных заболеваний. Сердечно-сосудистая система страдает от хронической симпатической гиперактивации: устойчивое повышение частоты сердечных сокращений и артериального давления ведёт к гипертрофии миокарда, ремоделированию сосудов, прогрессированию атеросклероза, повышенному риску аритмий и внезапной сердечной смерти. Желудочно-кишечный тракт, функция которого критически зависит от парасимпатической регуляции, страдает от её подавления: нарушается моторика, секреция, кровоснабжение слизистой, повышается проницаемость кишечного барьера, нарушается микробиом. Иммунная система модулируется вегетативными влияниями: хроническая симпатическая активация способствует провоспалительному сдвигу и одновременному подавлению противоинфекционного иммунитета. Метаболические последствия включают инсулинорезистентность, дислипидемию, накопление висцерального жира. Понимание этих механизмов объясняет, почему вмешательства, направленные на восстановление вегетативного баланса — релаксационные техники, дыхательные практики, стимуляция блуждающего нерва, физическая активность, — оказывают благотворное влияние на столь широкий спектр стресс-ассоциированных состояний.
3.3 Эндокринная система — медленные, но мощные сигналы
Эндокринная система обеспечивает второй основной канал регуляции стресс-ответа, принципиально отличающийся от нервной системы по временны́м параметрам, пространственному охвату и характеру воздействия. Если нервная система подобна электрической проводке здания, обеспечивающей мгновенную активацию конкретных приборов по определённым линиям, то эндокринная система подобна системе водоснабжения, медленно, но неуклонно доставляющей своё содержимое ко всем точкам подключения одновременно. Гормоны — сигнальные молекулы эндокринной системы — секретируются железами внутренней секреции непосредственно в кровоток и разносятся по всему организму, потенциально достигая каждой клетки. Однако эффект гормона реализуется только в клетках, экспрессирующих соответствующие рецепторы, что обеспечивает специфичность ответа при системном распространении сигнала. Временной масштаб эндокринной регуляции измеряется минутами, часами и днями, в отличие от миллисекундной скорости нервной передачи. Это означает, что эндокринные эффекты развиваются медленнее, но и сохраняются значительно дольше, обеспечивая устойчивую перестройку метаболизма и функций организма на период действия стрессора и на время восстановления после него. Эндокринная система обеспечивает долговременную адаптацию там, где нервная система обеспечивает немедленную реакцию.
Анатомически эндокринная система представлена совокупностью желез внутренней секреции, рассредоточенных по всему организму, но функционально интегрированных через гипоталамо-гипофизарный комплекс. Гипоталамус, небольшая область мозга, расположенная в основании промежуточного мозга, занимает уникальное положение на границе нервной и эндокринной систем: он получает нервные входы от многочисленных областей мозга, интегрирующих сенсорную информацию, эмоциональные оценки и когнитивные процессы, и преобразует эти нервные сигналы в эндокринные команды через продукцию рилизинг-гормонов и ингибирующих гормонов. Эти пептиды через специализированную портальную сосудистую систему достигают гипофиза — небольшой железы, расположенной в костном углублении основания черепа и называемой иногда «дирижёром эндокринного оркестра». Гипофиз, в свою очередь, секретирует тропные гормоны, регулирующие активность периферических эндокринных желез — надпочечников, щитовидной железы, половых желез. Эта иерархическая организация обеспечивает многоуровневый контроль эндокринных функций и создаёт возможности для тонкой регуляции через механизмы обратной связи. Помимо классических эндокринных желез, в продукции гормонов участвуют также сердце, почки, жировая ткань, желудочно-кишечный тракт и другие органы, что расширяет понимание эндокринной системы за пределы её традиционных анатомических границ.
Гипоталамо-гипофизарно-надпочечниковая ось занимает центральное место в эндокринной регуляции стресс-ответа и представляет собой одну из наиболее детально изученных нейроэндокринных систем. Активация этой оси начинается в гипоталамусе, где нейроны паравентрикулярного ядра синтезируют и высвобождают кортикотропин-рилизинг-гормон и аргинин-вазопрессин в портальную циркуляцию, связывающую гипоталамус с передней долей гипофиза. Эти пептиды стимулируют кортикотрофные клетки гипофиза к синтезу и секреции адренокортикотропного гормона, который поступает в системный кровоток и достигает коры надпочечников. Клетки пучковой зоны коры надпочечников в ответ на адренокортикотропный гормон усиливают синтез и секрецию глюкокортикоидов — у человека преимущественно кортизола. Весь каскад от восприятия стрессора до появления кортизола в крови занимает от нескольких минут до получаса, что значительно медленнее симпатоадреналовой реакции, но обеспечивает устойчивый ответ, сохраняющийся на протяжении часов. Пик концентрации кортизола достигается примерно через двадцать-тридцать минут после острого стресса, а возвращение к исходному уровню может занимать от одного до нескольких часов в зависимости от интенсивности и продолжительности стрессора.
Глюкокортикоиды обладают поразительно широким спектром биологических эффектов, затрагивающих метаболизм, иммунитет, функцию мозга, сердечно-сосудистую систему и практически все другие аспекты физиологии. Название «глюкокортикоиды» отражает их первоначально описанную роль в регуляции метаболизма глюкозы: они стимулируют глюконеогенез в печени, снижают поглощение глюкозы периферическими тканями, способствуют мобилизации аминокислот из мышц и жирных кислот из жировой ткани. Всё это направлено на обеспечение энергетическими субстратами мозга и работающих мышц в условиях стресса. Иммуномодулирующие эффекты глюкокортикоидов также были признаны давно: в фармакологических дозах они оказывают мощное противовоспалительное и иммуносупрессивное действие, что используется терапевтически при аутоиммунных и воспалительных заболеваниях. Физиологическое значение этих эффектов состоит в ограничении избыточного воспаления, которое может быть активировано стрессом, и в предотвращении аутоагрессии. Влияние глюкокортикоидов на мозг опосредовано высокой плотностью их рецепторов в гиппокампе, миндалине, префронтальной коре и включает модуляцию памяти, эмоций, когнитивных функций. Эти центральные эффекты имеют как адаптивное значение — усиление запоминания эмоционально значимых событий, — так и патогенное при хронической избыточности — нарушения памяти, настроения, когнитивный дефицит.
Метафора сравнения нервной и эндокринной коммуникации с различными формами человеческого общения помогает интуитивно понять их функциональные различия. Нервная система подобна экстренным текстовым сообщениям: мгновенная доставка, адресация конкретному получателю, краткое содержание, быстрое прочтение и удаление. Эндокринная система подобна длительным телефонным разговорам или, ещё точнее, рассылаемым по многим адресам циркулярным письмам: передача занимает время, сообщение получают многие адресаты одновременно, содержание требует осмысления и вызывает долгосрочные изменения в поведении получателей. Продолжая метафору, можно сказать, что острый стресс — это экстренное сообщение, требующее немедленной реакции; хронический стресс — это непрерывный поток писем, постепенно изменяющий всю систему приоритетов и распределения ресурсов. Понимание этих различий объясняет, почему немедленные эффекты стресса преимущественно определяются нервной системой, тогда как кумулятивные долгосрочные последствия — эндокринной.
Отрицательная обратная связь представляет собой ключевой механизм регуляции гипоталамо-гипофизарно-надпочечниковой оси, обеспечивающий самоограничение стресс-ответа и предотвращающий избыточное воздействие глюкокортикоидов на ткани. Кортизол, достигая высоких концентраций в крови, связывается с глюкокортикоидными рецепторами в гипоталамусе и гипофизе, что приводит к подавлению секреции кортикотропин-рилизинг-гормона и адренокортикотропного гормона соответственно. В результате стимуляция надпочечников ослабевает, продукция кортизола снижается, и система возвращается к исходному уровню после устранения стрессора. Этот механизм работает на нескольких временных масштабах: быстрая обратная связь реализуется в течение минут через негеномные эффекты глюкокортикоидов; отсроченная обратная связь развивается в течение часов через изменение экспрессии генов. Нарушение отрицательной обратной связи является одним из ключевых патогенетических механизмов при хроническом стрессе: рецепторы к глюкокортикоидам подвергаются даунрегуляции, чувствительность к тормозящему влиянию кортизола снижается, и ось остаётся хронически активированной даже при высоком уровне гормона. Этот феномен глюкокортикоидной резистентности связывает эндокринную дисрегуляцию с хроническим воспалением и другими патологическими последствиями длительного стресса.
Помимо гипоталамо-гипофизарно-надпочечниковой оси, стресс затрагивает и другие эндокринные системы, что вносит вклад в разнообразие его физиологических и патологических последствий. Симпатоадреналовая система, упомянутая ранее в контексте вегетативной регуляции, включает эндокринный компонент — мозговое вещество надпочечников, которое секретирует адреналин и норадреналин непосредственно в кровоток. Эти катехоламины действуют быстрее, чем глюкокортикоиды, но их эффект также более кратковременен. Гипоталамо-гипофизарно-тиреоидная ось при остром стрессе обычно подавляется, что может отражать адаптивное снижение базового метаболизма для перераспределения ресурсов; хронический стресс ассоциирован с нарушениями функции щитовидной железы. Гипоталамо-гипофизарно-гонадная ось чувствительна к стрессу: кортизол и кортикотропин-рилизинг-гормон подавляют секрецию гонадотропин-рилизинг-гормона, что ведёт к снижению продукции половых гормонов и нарушениям репродуктивной функции — аменорее у женщин, снижению либидо и сперматогенеза у мужчин. Гормон роста, пролактин, окситоцин, вазопрессин и множество других гормонов также вовлечены в стресс-ответ, создавая сложную картину эндокринных изменений.
Циркадная ритмичность эндокринной секреции представляет собой важный аспект нормальной физиологии, который существенно нарушается при хроническом стрессе. Секреция кортизола у здорового человека демонстрирует выраженный суточный ритм с максимумом в утренние часы вскоре после пробуждения — так называемый кортизоловый ответ на пробуждение, — постепенным снижением в течение дня и минимумом в первую половину ночи. Этот ритм синхронизирован с циклом сон-бодрствование и обеспечивает оптимальное распределение энергетических ресурсов: мобилизацию утром, когда начинается активная деятельность, и восстановление ночью. Хронический стресс нарушает этот ритм различными способами: уплощение суточной кривой кортизола, притупление утреннего пика, повышение вечерних и ночных уровней. Эти нарушения ритма сами по себе имеют патогенное значение независимо от среднего уровня гормона: они нарушают восстановительные процессы во время сна, десинхронизируют периферические тканевые часы, нарушают иммунную функцию. Восстановление нормальной циркадной ритмичности рассматривается как важная терапевтическая цель при стресс-связанных расстройствах, достижимая через нормализацию режима сна-бодрствования, световую терапию и другие хронотерапевтические вмешательства.
Понимание эндокринной регуляции стресса имеет прямые клинические и терапевтические следствия, которые выходят далеко за пределы эндокринологии в узком смысле. Измерение кортизола и других гормонов стресса используется для оценки функционального состояния гипоталамо-гипофизарно-надпочечниковой оси, хотя интерпретация результатов требует учёта циркадного ритма, характера стрессора, индивидуальных особенностей реактивности. Волосяной кортизол как интегральный показатель секреции за месяцы приобретает растущее значение для оценки хронического стресса. Фармакологическая модуляция эндокринных систем применяется при ряде состояний: заместительная терапия при надпочечниковой недостаточности, подавление избыточной продукции при синдроме Кушинга, модуляция рецепторов при разработке новых психотропных препаратов. Однако не менее важны нефармакологические подходы: психотерапия, изменяющая когнитивную оценку стрессоров, может нормализовать активность гипоталамо-гипофизарно-надпочечниковой оси; физическая активность модулирует эндокринный ответ на стресс; социальная поддержка снижает кортизоловую реактивность. Интеграция понимания эндокринных механизмов с психологическими и поведенческими вмешательствами представляет собой сущность современного подхода к терапии стресс-связанных состояний.
3.4 Иммунная система — не только защита, но и регуляция
Традиционное представление об иммунной системе как об автономной защитной структуре, единственной функцией которой является распознавание и уничтожение патогенов, претерпело радикальную трансформацию в свете открытий последних десятилетий. Современная иммунология рассматривает иммунную систему не просто как «армию» организма, ведущую войну с внешними захватчиками, но как сложнейшую регуляторную сеть, интегрированную с нервной и эндокринной системами и участвующую в поддержании гомеостаза, восстановлении тканей, модуляции метаболизма, регуляции поведения и когнитивных функций. Эта расширенная концепция иммунитета имеет фундаментальное значение для понимания стресса, поскольку именно через иммунные механизмы реализуются многие связи между психологическими переживаниями и физиологическими изменениями, между субъективным опытом и объективными показателями здоровья. Иммунная система оказалась тем самым звеном, которое связывает душу и тело не в метафорическом, а в буквальном молекулярном смысле: сигнальные молекулы, продуцируемые иммунными клетками, способны достигать мозга и изменять его функцию, а сигналы из мозга, в свою очередь, модулируют иммунные реакции. Признание этой двунаправленной связи породило новую научную дисциплину — психонейроиммунологию, — которая изучает взаимодействия между психикой, нервной системой и иммунитетом и открывает новые перспективы для понимания и лечения стресс-связанных заболеваний.
Цитокины — сигнальные белки, секретируемые иммунными клетками — представляют собой молекулярных посредников, через которых иммунная система осуществляет свою регуляторную функцию, влияя на работу практически всех органов и систем, включая мозг. Семейство цитокинов насчитывает сотни различных молекул, включающих интерлейкины, интерфероны, факторы некроза опухоли, хемокины, факторы роста и другие. Каждый цитокин имеет свой спектр действия, свои клетки-мишени и свой профиль эффектов, однако в контексте стресса особое значение имеют провоспалительные цитокины — интерлейкин-один-бета, интерлейкин-шесть и фактор некроза опухоли альфа. Эти молекулы продуцируются активированными макрофагами, моноцитами, дендритными клетками и другими клетками врождённого иммунитета в ответ на распознавание патогенов или сигналов повреждения тканей. Однако, как оказалось, те же цитокины продуцируются и при отсутствии инфекции — в ответ на психологический стресс, что опосредовано влиянием симпатической нервной системы и гормонов стресса на иммунные клетки. Провоспалительные цитокины способны достигать мозга несколькими путями: через области с проницаемым гематоэнцефалическим барьером, через активный транспорт специализированными переносчиками, через афферентные волокна блуждающего нерва, через индукцию вторичных медиаторов в эндотелии мозговых сосудов. Достигнув мозга, цитокины оказывают глубокое влияние на нейромедиаторный метаболизм, нейроэндокринную регуляцию и поведение.
Влияние цитокинов на мозг реализуется через комплекс нейрохимических изменений, которые лежат в основе характерных психологических и поведенческих проявлений иммунной активации. Провоспалительные цитокины активируют фермент индоламин-два-три-диоксигеназу, который превращает триптофан — предшественник серотонина — в кинуренин, направляя метаболизм по альтернативному пути и снижая доступность субстрата для синтеза серотонина. Это частично объясняет связь между воспалением и депрессией, поскольку серотониновая недостаточность традиционно рассматривается как один из механизмов депрессивных расстройств. Одновременно цитокины влияют на дофаминергическую передачу, снижая активность дофаминовой системы в области вознаграждения, что может лежать в основе ангедонии — утраты способности испытывать удовольствие, — являющейся ключевым симптомом депрессии. Глутаматергическая и ГАМКергическая передача также модулируются провоспалительными сигналами, нарушая баланс возбуждения и торможения в нейронных сетях. Кроме того, цитокины активируют гипоталамо-гипофизарно-надпочечниковую ось, повышая продукцию кортизола, и могут нарушать механизмы глюкокортикоидной обратной связи, способствуя хронической дисрегуляции оси. Все эти нейрохимические изменения суммируются в характерный клинический синдром, известный как «болезненное поведение».
Болезненное поведение представляет собой организованный поведенческий ответ на иммунную активацию, который эволюционно сформировался как адаптивная стратегия сохранения энергии для борьбы с инфекцией. Этот синдром включает усталость и сонливость, снижение двигательной активности, потерю аппетита, социальную изоляцию, снижение интереса к окружающему миру, когнитивные нарушения, подавленное настроение. Перечисленные проявления поразительно напоминают симптомы клинической депрессии, что неслучайно: они опосредованы сходными нейрохимическими механизмами. В эволюционном контексте болезненное поведение адаптивно: когда организм борется с инфекцией, энергетически затратная социальная и исследовательская активность должна быть минимизирована, чтобы ресурсы могли быть направлены на иммунный ответ. Социальная изоляция также снижает риск передачи инфекции другим членам группы, что имеет значение для выживания вида. Однако при хроническом стрессе, когда провоспалительная активация персистирует в отсутствие реальной инфекции, болезненное поведение утрачивает адаптивный смысл и превращается в источник страдания, феноменологически неотличимого от депрессии. Эта концепция послужила основой для воспалительной гипотезы депрессии, которая открыла новые терапевтические перспективы.
Влияние стресса на иммунную функцию носит двунаправленный и зависящий от временно́й динамики характер, что объясняет кажущуюся противоречивость данных о связи стресса и иммунитета. Острый стресс, продолжительностью от минут до нескольких часов, как правило, оказывает иммуностимулирующее действие, что функционально адаптивно: если организм сталкивается с угрозой, которая может повлечь травму и инфицирование, усиление иммунной готовности повышает шансы на выживание. При остром стрессе наблюдается перераспределение иммунных клеток из депо — селезёнки, костного мозга, лимфатических узлов — в кровоток и далее в ткани, наиболее подверженные риску повреждения: кожу, слизистые оболочки, подкожную клетчатку. Функциональная активность иммунных клеток при остром стрессе также может повышаться: усиливается цитотоксичность естественных киллеров, фагоцитарная активность макрофагов, продукция антител. Классические эксперименты демонстрируют, что умеренный острый стресс непосредственно перед вакцинацией может усиливать последующий иммунный ответ. Однако эта иммуностимуляция ограничена во времени: если стресс продолжается или повторяется, иммуностимулирующие эффекты сменяются противоположными.
Хронический стресс оказывает преимущественно иммуносупрессивное влияние на адаптивный иммунитет, одновременно способствуя развитию хронического воспаления низкой интенсивности — парадоксальная комбинация, имеющая серьёзные последствия для здоровья. Подавление адаптивного иммунитета при хроническом стрессе проявляется снижением пролиферативного ответа лимфоцитов на митогены, ослаблением продукции антител, угнетением цитотоксической активности, нарушением формирования иммунологической памяти. Эти изменения опосредованы хроническим воздействием глюкокортикоидов, которые в высоких концентрациях оказывают выраженное иммуносупрессивное действие на лимфоциты. Клинически это проявляется повышенной восприимчивостью к инфекциям: люди, находящиеся в состоянии хронического стресса — ухаживающие за больными родственниками, переживающие хроническое одиночество или социальную изоляцию, сталкивающиеся с продолжительными профессиональными или финансовыми трудностями, — демонстрируют более частые и тяжёлые простудные заболевания, более медленное заживление ран, более слабый ответ на вакцинацию. Однако одновременно с подавлением противоинфекционного иммунитета при хроническом стрессе активируется другая ветвь иммунного ответа — системное воспаление.
Системное воспаление низкой интенсивности при хроническом стрессе характеризуется умеренным, но устойчивым повышением уровней провоспалительных цитокинов и белков острой фазы воспаления в крови при отсутствии инфекции или очевидного повреждения тканей. В отличие от острого воспаления, которое является локализованным, интенсивным, самоограничивающимся ответом на конкретную угрозу, хроническое воспаление низкой интенсивности персистирует неопределённо долго и оказывает повреждающее действие на ткани по всему организму. Повышенные уровни С-реактивного белка, интерлейкина-шесть, фибриногена и других маркёров воспаления обнаруживаются у людей с хроническим стрессом даже при контроле очевидных воспалительных заболеваний. Это «тлеющее» воспаление не вызывает классических симптомов острого воспаления — покраснения, отёка, боли, повышения температуры, — но вносит вклад в патогенез широкого спектра хронических заболеваний: атеросклероза и сердечно-сосудистой патологии, метаболического синдрома и диабета второго типа, нейродегенерации, депрессии и других психических расстройств. Механизмы, через которые хронический стресс индуцирует воспаление, включают нарушение глюкокортикоидной супрессии воспалительного ответа (глюкокортикоидную резистентность), повышенное высвобождение провоспалительных сигналов из жировой ткани, нарушение барьерной функции кишечника с транслокацией бактериальных компонентов.
Микроглия — резидентные иммунные клетки центральной нервной системы — занимает особое место в контексте нейроиммунных взаимодействий при стрессе и заслуживает отдельного рассмотрения. В отличие от большинства иммунных клеток, которые происходят из костномозговых предшественников и населяют периферические ткани, микроглия имеет особое эмбриональное происхождение из желточного мешка и заселяет мозг на ранних этапах развития, оставаясь там на протяжении всей жизни. В здоровом мозге микроглия находится в так называемом «наблюдательном» состоянии, непрерывно мониторируя микроокружение, удаляя клеточный дебрис и участвуя в ремоделировании синапсов — процессе, необходимом для нормального обучения и памяти. Однако при воспалительных сигналах — как периферических, достигающих мозга через описанные выше пути, так и центральных — микроглия активируется, переходя в провоспалительное состояние с характерными морфологическими и функциональными изменениями. Активированная микроглия продуцирует провоспалительные цитокины, хемокины, активные формы кислорода и азота, которые могут повреждать нейроны и нарушать синаптическую функцию. Исследования на животных моделях демонстрируют, что хронический стресс вызывает устойчивую активацию микроглии в гиппокампе, префронтальной коре, миндалине и других областях мозга, связанных с эмоциями и когнитивными функциями.
Концепция нейровоспаления приобретает всё большее значение для понимания патогенеза стресс-связанных нарушений мозга, включая депрессию, тревожные расстройства, когнитивные нарушения. Нейровоспаление характеризуется активацией микроглии и астроцитов, локальной продукцией провоспалительных медиаторов, нарушением гематоэнцефалического барьера, инфильтрацией периферическими иммунными клетками в тяжёлых случаях. В отличие от классического периферического воспаления, нейровоспаление имеет свои особенности, обусловленные уникальной иммунологией мозга: отсутствием классического лимфатического дренажа (хотя недавно открытые менингеальные лимфатические сосуды пересматривают это представление), ограниченным проникновением периферических иммунных клеток через гематоэнцефалический барьер, преобладанием резидентных микроглиальных клеток. Постмортемные исследования мозга людей с депрессией выявляют признаки микроглиальной активации, а позитронно-эмиссионная томография с лигандами к маркёрам активированной микроглии демонстрирует усиленный сигнал у пациентов с депрессией по сравнению со здоровыми добровольцами. Эти данные поддерживают представление о нейровоспалении как о важном патогенетическом механизме при стресс-связанных психических расстройствах и открывают перспективы для противовоспалительных терапевтических подходов.
Терапевтические импликации понимания иммунной регуляции при стрессе включают как традиционные подходы в новом свете, так и инновационные стратегии. Противовоспалительная терапия при депрессии исследуется в многочисленных клинических испытаниях: нестероидные противовоспалительные препараты, ингибиторы циклооксигеназы-два, антицитокиновые препараты демонстрируют антидепрессивные эффекты в ряде исследований, особенно у пациентов с повышенными маркёрами воспаления. Это открывает путь к персонализированному подходу, где выбор терапии основывается на иммунном профиле пациента. Традиционные антидепрессанты, как оказалось, обладают противовоспалительными свойствами, что может вносить вклад в их терапевтическое действие. Психотерапия также модулирует иммунные параметры: успешная когнитивно-поведенческая терапия ассоциирована со снижением маркёров воспаления. Образ жизни — физическая активность, диета, сон, социальные связи — оказывает мощное влияние на иммунную функцию и воспалительный статус, что обосновывает включение этих компонентов в комплексное лечение стресс-связанных расстройств. Понимание иммунной регуляции при стрессе не просто добавляет ещё один уровень сложности, но открывает новые терапевтические окна и объясняет механизмы уже существующих эффективных вмешательств.
3.5 Другие системы организма под влиянием стресса
Три главные регуляторные системы — нервная, эндокринная и иммунная — представляют собой командные центры стресс-ответа, однако исполнителями этих команд являются все остальные системы организма, каждая из которых вносит свой вклад в адаптацию и несёт свою долю последствий при хронической перегрузке. Понимание того, как стресс затрагивает сердечно-сосудистую, дыхательную, пищеварительную, мочеполовую, опорно-двигательную и другие системы, необходимо для осмысления полного спектра физиологических и клинических проявлений стресса. Общий принцип, объединяющий изменения во всех этих системах, состоит в перераспределении ресурсов: организм, воспринимающий угрозу, направляет энергию и кровоснабжение к системам, критически важным для немедленного выживания — мышцам, сердцу, мозгу, — за счёт систем, обеспечивающих долгосрочное функционирование — пищеварения, репродукции, роста, восстановления. Это перераспределение адаптивно при кратковременном стрессе, но становится источником патологии при хроническом, поскольку постоянно «обделённые» системы накапливают нарушения. Кроме того, хронические изменения в каждой из этих систем посылают обратные сигналы к регуляторным центрам, замыкая порочные круги, которые поддерживают и усугубляют дисрегуляцию.
Сердечно-сосудистая система является одной из наиболее непосредственных мишеней стресс-ответа и одновременно наиболее хорошо изученной с точки зрения связи со стрессом. Острый стресс вызывает характерный паттерн сердечно-сосудистых изменений: учащение сердечных сокращений под влиянием симпатической активации и снижения вагального тонуса, усиление сократимости миокарда, сужение периферических сосудов с перераспределением крови к мышцам, повышение артериального давления. Эти изменения обеспечивают готовность к интенсивной физической активности и быстро нормализуются после устранения угрозы. Однако при хроническом стрессе постоянная симпатическая активация приводит к устойчивой гемодинамической нагрузке, которая влечёт за собой структурные и функциональные изменения. Хроническое повышение артериального давления вызывает гипертрофию миокарда левого желудочка — компенсаторную адаптацию, которая со временем переходит в патологическое ремоделирование с нарушением диастолической функции. Эндотелий сосудов, постоянно подвергающийся воздействию катехоламинов и кортизола, утрачивает нормальную функцию вазодилатации и приобретает провоспалительный, протромботический фенотип. Хроническое воспаление низкой интенсивности ускоряет атерогенез — формирование атеросклеротических бляшек в артериях. Вариабельность сердечного ритма снижается, что отражает нарушение вегетативной регуляции и само по себе является предиктором сердечно-сосудистой смертности.
Дыхательная система непосредственно вовлекается в острый стресс-ответ и может становиться источником проблем при хроническом стрессе. При остром стрессе дыхание учащается и углубляется, обеспечивая повышенное поступление кислорода и выведение углекислого газа в соответствии с возросшими метаболическими потребностями. Бронхи расширяются под влиянием симпатической активации и адреналина, снижая сопротивление воздушному потоку. Эти изменения адаптивны для кратковременного физического напряжения, однако при хроническом стрессе паттерн дыхания может устойчиво изменяться. Гипервентиляция — избыточное дыхание, не соответствующее метаболическим потребностям — является частым спутником тревожных состояний и ведёт к гипокапнии (снижению уровня углекислого газа в крови), что вызывает сужение мозговых сосудов, головокружение, парестезии, усиливающие тревогу и замыкающие порочный круг. У пациентов с бронхиальной астмой психологический стресс может провоцировать бронхоспазм и обострения заболевания, что опосредовано как прямыми нервными влияниями, так и иммунными механизмами. Исследования демонстрируют связь между хроническим стрессом и ухудшением течения хронических респираторных заболеваний, а также повышенным риском респираторных инфекций вследствие иммуносупрессии.
Пищеварительная система особенно чувствительна к стрессу благодаря богатой вегетативной иннервации и наличию собственной «кишечной» нервной системы, содержащей больше нейронов, чем спинной мозг. При остром стрессе пищеварительная функция подавляется: снижается секреция слюны (сухость во рту — типичный симптом тревоги), замедляется моторика желудка и кишечника, уменьшается кровоснабжение желудочно-кишечного тракта, которое перенаправляется к мышцам. Это адаптивно: в ситуации угрозы переваривание пищи может подождать. Однако при хроническом стрессе пищеварительная система страдает от постоянного подавления и нарушения регуляции. Функциональные желудочно-кишечные расстройства — синдром раздражённого кишечника, функциональная диспепсия — тесно связаны со стрессом и рассматриваются как нарушения оси «мозг-кишечник». Стресс повышает проницаемость кишечного барьера — феномен «протекающего кишечника», — что позволяет бактериальным компонентам проникать в кровоток и поддерживать системное воспаление. Микробиом кишечника, насчитывающий триллионы бактерий и играющий критическую роль в метаболизме, иммунитете и даже функции мозга, изменяется под влиянием хронического стресса в сторону дисбиоза — нарушения нормального состава и функции микробного сообщества.
Репродуктивная система является одной из первых «жертв» перераспределения ресурсов при хроническом стрессе, что имеет эволюционную логику: в условиях угрозы выживанию размножение может и должно быть отложено. Кортикотропин-рилизинг-гормон и глюкокортикоиды подавляют секрецию гонадотропин-рилизинг-гормона в гипоталамусе, что ведёт к снижению продукции лютеинизирующего и фолликулостимулирующего гормонов гипофизом и, соответственно, к угнетению функции половых желез. У женщин хронический стресс может вызывать нарушения менструального цикла вплоть до аменореи, ановуляцию, снижение фертильности. У мужчин наблюдается снижение уровня тестостерона, нарушения сперматогенеза, эректильная дисфункция. Стресс во время беременности ассоциирован с повышенным риском преждевременных родов, низкой массы тела новорождённого и, что особенно важно, с долгосрочными последствиями для развивающегося плода — так называемым пренатальным программированием, которое влияет на стресс-реактивность, метаболизм и риск заболеваний на протяжении всей жизни потомства. Эти данные подчёркивают межпоколенческие последствия хронического стресса и важность его профилактики.
Опорно-двигательная система вовлекается в стресс-ответ через механизмы мышечного напряжения, которые хорошо знакомы каждому, кто когда-либо испытывал стресс. При остром стрессе скелетные мышцы напрягаются, подготавливаясь к действию — бегству или борьбе. Это напряжение особенно выражено в мышцах шеи, плечевого пояса, спины, лица. При хроническом стрессе мышечное напряжение становится устойчивым, приводя к дискомфорту, боли, ограничению подвижности. Головные боли напряжения — наиболее распространённый тип головных болей — непосредственно связаны с хроническим напряжением перикраниальных мышц. Боли в шее и спине, эпидемически распространённые в современном обществе, имеют стрессовый компонент в своей этиологии. Височно-нижнечелюстная дисфункция, проявляющаяся болью в области челюстного сустава и затруднением жевания, связана с бруксизмом — непроизвольным сжатием и скрежетанием зубами, усиливающимся при стрессе. Хроническое мышечное напряжение не только причиняет дискомфорт само по себе, но и может формировать патологические постуральные паттерны, нарушать кровообращение в напряжённых мышцах, создавать триггерные точки и способствовать развитию миофасциального болевого синдрома.
Кожа, являясь крупнейшим органом тела и барьером между организмом и внешней средой, также демонстрирует выраженные реакции на стресс. Острый стресс вызывает бледность кожи вследствие сужения кожных сосудов и перераспределения крови к мышцам, усиление потоотделения (особенно на ладонях и подошвах — эмоциональное потоотделение), появление «гусиной кожи» вследствие сокращения мышц, поднимающих волосы. При хроническом стрессе страдают барьерная функция кожи, заживление ран, иммунная защита. Многие дерматологические заболевания обостряются при стрессе или имеют стрессовый компонент в своей этиологии: псориаз, атопический дерматит, акне, розацеа, алопеция (выпадение волос), витилиго. Термин «психодерматология» обозначает область, изучающую связи между психикой и кожными заболеваниями. Кожа богато иннервирована и содержит собственные иммунные клетки, что обеспечивает её чувствительность к нейроэндокринным и воспалительным сигналам стресса. Старение кожи ускоряется при хроническом стрессе через механизмы окислительного повреждения, укорочения теломер, накопления сенесцентных клеток — те же механизмы, которые опосредуют системное ускоренное старение при стрессе.
Выделительная система, включающая почки и мочевыводящие пути, также подвержена влиянию стресса, хотя эти эффекты менее изучены, чем изменения в других системах. При остром стрессе почечный кровоток снижается вследствие симпатической вазоконстрикции, что может временно уменьшать диурез. Антидиуретический гормон (вазопрессин), секреция которого усиливается при стрессе, способствует задержке воды. Хронический стресс ассоциирован с повышенным риском хронической болезни почек, хотя механизмы этой связи сложны и включают влияние гипертензии, метаболических нарушений, воспаления. Функциональные нарушения мочеиспускания — учащённое мочеиспускание, императивные позывы, синдром раздражённого мочевого пузыря — часто связаны со стрессом и тревогой. Эти проявления объясняются влиянием вегетативной нервной системы на тонус мочевого пузыря и сфинктеров, а также центральными механизмами регуляции мочеиспускания, чувствительными к эмоциональному состоянию. Интерстициальный цистит — хронический болевой синдром мочевого пузыря — рассматривается некоторыми исследователями как стресс-чувствительное расстройство, сходное с синдромом раздражённого кишечника.
Интеграция знаний о влиянии стресса на различные системы организма создаёт картину системной перестройки, в которой ни один орган не остаётся незатронутым. Эта системность имеет несколько важных следствий. Во-первых, она объясняет полисимптомность стресс-связанных состояний: пациент с хроническим стрессом может одновременно жаловаться на головные боли, нарушения пищеварения, сердцебиения, кожные проблемы, и все эти жалобы имеют единую этиологическую основу. Во-вторых, она указывает на необходимость холистического подхода к диагностике и лечению, учитывающего взаимосвязи между системами. В-третьих, она подчёркивает потенциал системных вмешательств: если стресс вызывает системные изменения, то и снижение стресса может приносить системные улучшения. Образ жизни, управление стрессом, психотерапия, социальная поддержка влияют не на один орган, а на весь организм как целое, что объясняет широту их терапевтических эффектов.
3.6 Интеграция систем — организм как единое целое
Рассмотрение отдельных систем организма, при всей его аналитической ценности, создаёт риск утраты видения целого — того интегрированного единства, которым является живой организм. Тело не является механической суммой своих частей; оно представляет собой сложную динамическую систему, в которой все компоненты непрерывно взаимодействуют, модулируя функции друг друга и порождая эмерджентные свойства, несводимые к свойствам отдельных элементов. Стресс-ответ является превосходной иллюстрацией этой интеграции: он не локализован ни в одной системе, но представляет собой скоординированную перестройку всего организма, вовлекающую нервные, эндокринные, иммунные, метаболические, поведенческие и психологические компоненты в единый адаптивный ответ. Понимание механизмов этой интеграции и последствий её нарушения составляет одну из центральных задач науки о стрессе. Метафора оркестра, введённая ранее, может быть развита для осмысления интегративных аспектов: если отдельные системы подобны секциям оркестра, то интеграция — это партитура и дирижёр, обеспечивающие согласованное исполнение единого произведения. Когда интеграция нарушается, отдельные инструменты начинают играть каждый свою мелодию, порождая какофонию, которая и представляет собой патологию хронического стресса.
Механизмы интеграции между системами многообразны и включают нервные, гуморальные и молекулярные каналы коммуникации. Нервная интеграция обеспечивается анатомическими связями между различными уровнями мозга и между центральной нервной системой и периферическими органами. Гипоталамус занимает ключевое положение в этой архитектуре, получая входы от коры, лимбических структур, ствола мозга, внутренних органов и интегрируя эту информацию для координации вегетативных, эндокринных и поведенческих ответов. Гуморальная интеграция осуществляется через гормоны и цитокины, циркулирующие в крови и достигающие всех тканей одновременно. Кортизол, например, влияет практически на все клетки организма благодаря повсеместному распределению глюкокортикоидных рецепторов. Молекулярная интеграция реализуется через общие сигнальные пути и транскрипционные факторы, которые координируют ответы на уровне отдельных клеток. Транскрипционный фактор ядерный фактор каппа-В активируется как воспалительными, так и стрессовыми сигналами и координирует экспрессию сотен генов, участвующих в иммунном ответе, метаболизме, выживании клеток. Эти многоуровневые механизмы интеграции обеспечивают согласованность организменного ответа на стресс.
Обратная связь представляет собой фундаментальный принцип регуляции, обеспечивающий самоконтроль и самокоррекцию физиологических систем. Отрицательная обратная связь — наиболее распространённый тип — работает по принципу термостата: когда регулируемый параметр отклоняется от заданного значения, активируются механизмы, возвращающие его к норме. В контексте стресса классическим примером является глюкокортикоидная обратная связь в гипоталамо-гипофизарно-надпочечниковой оси: повышение уровня кортизола подавляет секрецию кортикотропин-рилизинг-гормона и адренокортикотропного гормона, ограничивая дальнейшую продукцию кортизола и обеспечивая самолимитирование стресс-ответа. Аналогичные механизмы обратной связи действуют во всех регуляторных системах. Барорефлекс обеспечивает обратную связь в регуляции артериального давления: повышение давления активирует барорецепторы, которые через рефлекторную дугу вызывают снижение симпатического тонуса и повышение вагального, возвращая давление к норме. Воспалительный ответ самоограничивается через продукцию противовоспалительных медиаторов, включая глюкокортикоиды и противовоспалительные цитокины. Нарушение механизмов обратной связи является ключевым патогенетическим механизмом при хроническом стрессе.
Дисрегуляция при хроническом стрессе характеризуется нарушением нормальных петель обратной связи, что приводит к утрате гомеостатического контроля и персистенции патологических состояний. Глюкокортикоидная резистентность — снижение чувствительности тканей к тормозящему действию кортизола — является одним из наиболее изученных примеров такой дисрегуляции. При хроническом стрессе глюкокортикоидные рецепторы в гипоталамусе и гипофизе подвергаются даунрегуляции, что ослабляет отрицательную обратную связь и позволяет оси оставаться хронически активированной даже при высоких уровнях кортизола. Одновременно глюкокортикоидная резистентность иммунных клеток означает, что высокий кортизол не может эффективно подавлять воспаление, и провоспалительные процессы персистируют. Вегетативный дисбаланс с преобладанием симпатической активации и ослаблением парасимпатической сопровождается снижением барорефлекторной чувствительности, что нарушает нормальную регуляцию артериального давления. Нарушение циркадной ритмичности — уплощение суточных кривых кортизола, дезорганизация сна — представляет собой ещё одну форму дисрегуляции, нарушающую временну́ю координацию физиологических процессов. Все эти формы дисрегуляции взаимосвязаны и усиливают друг друга, создавая паттерн системной дезинтеграции.
Концепция аллостаза и аллостатической нагрузки, разработанная нейроэндокринологом Брюсом Макьюэном, предоставляет интегративную рамку для понимания кумулятивных последствий хронического стресса на системном уровне. Термин «аллостаз» буквально означает «достижение стабильности через изменение» и описывает процесс, посредством которого организм адаптируется к стрессорам путём активной перестройки физиологических параметров. В отличие от классического гомеостаза, предполагающего поддержание параметров в узком диапазоне вокруг фиксированных заданных значений, аллостаз признаёт, что заданные значения сами могут изменяться в зависимости от требований среды. Например, артериальное давление закономерно повышается при физической нагрузке — это не нарушение гомеостаза, а аллостатическая адаптация к изменившимся потребностям. Однако каждый эпизод аллостатической адаптации сопряжён с физиологическими издержками, и при повторяющемся или хроническом стрессе эти издержки накапливаются, формируя аллостатическую нагрузку — кумулятивный износ физиологических систем вследствие хронической адаптации к стрессорам. Аллостатическая нагрузка проявляется в виде повышенного артериального давления, абдоминального ожирения, нарушения регуляции глюкозы, дислипидемии, маркёров воспаления — комплекса изменений, предсказывающего повышенный риск сердечно-сосудистых заболеваний, диабета, когнитивного снижения и преждевременной смерти.
Системная дезинтеграция при хроническом стрессе может быть понята через метафору оркестра, в котором дирижёр утрачивает контроль. В здоровом организме мозг, и особенно его интегративные структуры — гипоталамус, лимбическая система, префронтальная кора, — выполняет функцию дирижёра, координирующего активность всех систем в соответствии с требованиями текущей ситуации и долгосрочными целями организма. Отдельные системы, подобно секциям оркестра, следуют этому руководству, вступая в нужный момент, играя с нужной интенсивностью, согласовывая свои партии друг с другом. При хроническом стрессе эта координация нарушается: симпатическая система продолжает «играть громко» даже когда требуется тишина, гипоталамо-гипофизарно-надпочечниковая ось не отвечает на сигналы к снижению громкости, иммунная система начинает свою собственную партию воспаления, не согласованную с остальным оркестром. Метаболические системы перестраиваются на режим постоянной мобилизации, несовместимый с требованиями повседневной жизни. Результатом является какофония — рассогласованность физиологических процессов, которая и представляет собой сущность хронического стресса на системном уровне.
Восстановление интеграции является ключевой терапевтической задачей при хроническом стрессе, и понимание механизмов интеграции указывает на потенциальные точки вмешательства. Воздействие на центральные интегративные структуры — через психотерапию, изменяющую когнитивные и эмоциональные паттерны, через медитативные практики, тренирующие внимание и эмоциональную регуляцию, через фармакологические агенты, модулирующие нейромедиаторные системы, — может восстанавливать нормальную «дирижёрскую» функцию мозга. Воздействие на механизмы обратной связи — через физическую активность, нормализующую глюкокортикоидную чувствительность, через дыхательные практики, повышающие барорефлекторную чувствительность и вагальный тонус, через противовоспалительные вмешательства, разрывающие порочные круги воспаления, — может восстанавливать нормальные регуляторные контуры. Воздействие на отдельные системы — через специфические фармакологические агенты, модифицирующие активность конкретных путей, — может корректировать локальные нарушения, хотя системный характер проблемы требует и системных решений. Образ жизни — сон, питание, физическая активность, социальные связи — оказывает комплексное воздействие на множество систем одновременно и представляет собой фундаментальную основу восстановления интеграции.
Понимание организма как интегрированного целого имеет глубокие философские и практические импликации для подхода к стрессу. Философски это преодолевает картезианский дуализм, разделявший ум и тело, душу и материю, и предлагает взамен холистическое видение, в котором психологическое и физиологическое представляют собой разные аспекты единой реальности. Субъективное переживание стресса и объективные физиологические изменения — не две отдельные сущности, а два измерения одного процесса, разворачивающегося одновременно на всех уровнях организации живой системы. Практически это означает, что эффективная помощь при стресс-связанных проблемах не может ограничиваться ни чисто биологическими вмешательствами, игнорирующими психологический контекст, ни чисто психологическими, игнорирующими телесную реальность. Интегративный подход, признающий единство ума и тела, психики и физиологии, индивидуума и социального контекста, представляет собой наиболее адекватный ответ на системную природу стресса. Последующие модули курса будут детализировать механизмы, обозначенные в этом вводном обзоре, однако интегративная перспектива, установленная здесь, должна сопровождать все дальнейшие погружения в частности, сохраняя видение целого за деревьями леса.
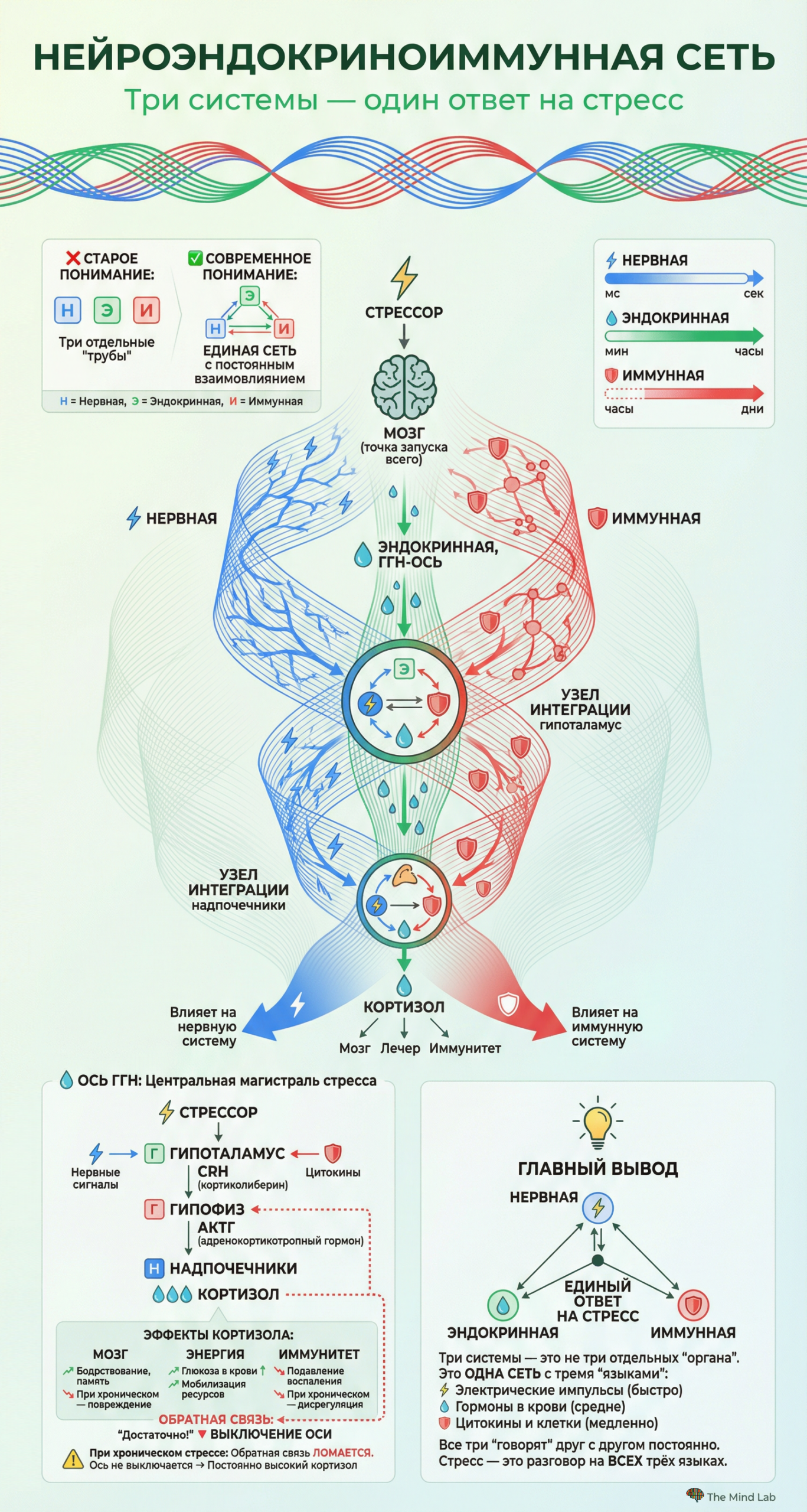
4. Психологический уровень — от биологии к субъективному опыту
4.1 Эмерджентность — как субъективное рождается из биологии
Переход от системного уровня анализа к психологическому представляет собой не просто очередной шаг в иерархии организации живой материи, но качественный скачок, порождающий феномены принципиально иного порядка. На молекулярном, клеточном и системном уровнях мы имели дело с объективными процессами, доступными измерению приборами и описанию на языке физики и химии: концентрации гормонов, частоты нейронных разрядов, паттерны иммунной активации. На психологическом уровне перед нами возникает нечто качественно иное — субъективный опыт, переживание, чувство, смысл. Когда человек говорит «мне страшно» или «я чувствую себя подавленным», он описывает не концентрацию кортизола в крови и не активность миндалины, хотя эти биологические процессы безусловно происходят в его организме. Он описывает свой внутренний мир — то, как ситуация переживается изнутри, каково это быть испуганным или подавленным. Этот субъективный, феноменологический аспект стресса не менее реален, чем его биологические корреляты, и именно он определяет, как человек осмысляет свою ситуацию и как действует в ней. Понимание отношений между биологическим и психологическим уровнями составляет одну из центральных философских проблем науки о стрессе и требует обращения к концепции эмерджентности как ключу к осмыслению этих отношений.
Концепция эмерджентности, или эмергентности, описывает фундаментальное свойство сложных систем порождать качества, которые не присутствуют в их отдельных компонентах и не могут быть предсказаны из свойств этих компонентов, рассматриваемых изолированно. Термин происходит от латинского слова, означающего «появляться», «возникать», и в философии науки обозначает появление новых свойств на более высоких уровнях организации материи. Классическим примером служит вода: молекула воды состоит из двух атомов водорода и одного атома кислорода, однако ни водород, ни кислород по отдельности не обладают свойством «мокрости» — способностью смачивать поверхности и вызывать характерное тактильное ощущение. Это свойство появляется только тогда, когда множество молекул воды взаимодействует друг с другом и с другими веществами, формируя макроскопический объект с характерными физическими свойствами. Аналогично, ни один отдельный нейрон не «думает», не «чувствует», не «принимает решений» — эти свойства возникают только на уровне мозга как целостной системы из миллиардов взаимодействующих нейронов. Сознание, эмоции, мышление являются эмерджентными свойствами нервной системы, несводимыми к свойствам её элементов, хотя и зависящими от них.
Признание эмерджентности психологических феноменов позволяет преодолеть две крайние позиции, каждая из которых по-своему искажает понимание стресса. Первая крайность — редукционизм, утверждающий, что психологические явления суть «просто» биология, «просто» химия мозга, и что, следовательно, описание на уровне нейромедиаторов и гормонов является полным и исчерпывающим. Эта позиция игнорирует качественную специфику психологического уровня: знание концентрации серотонина ничего не говорит нам о содержании мыслей, о смысле переживания, о значении события для конкретного человека. Вторая крайность — дуализм, постулирующий существование нематериальной психики, независимой от тела и лишь таинственным образом связанной с ним. Эта позиция несовместима с данными нейронауки, демонстрирующими тесную корреляцию между психологическими состояниями и активностью мозга, и с эволюционной биологией, показывающей постепенное развитие когнитивных способностей в животном царстве. Концепция эмерджентности предлагает третий путь: психологические феномены реальны, несводимы к биологии, но возникают из биологии и зависят от неё. Это позиция, иногда называемая нередуктивным физикализмом, позволяет признавать специфику психологического уровня без обращения к сверхъестественным сущностям.
Применение концепции эмерджентности к пониманию стресса имеет глубокие практические следствия, касающиеся как исследования, так и терапии. Исследовательское следствие состоит в признании того, что полное понимание стресса требует изучения на всех уровнях — от молекулярного до психологического и социального — и что данные каждого уровня несводимы к данным других уровней, хотя и связаны с ними. Измерение кортизола не заменяет психологическую оценку субъективного переживания стресса, равно как субъективные отчёты не заменяют биологических маркёров. Оба типа данных информативны и необходимы. Терапевтическое следствие состоит в признании множественных точек входа для интервенций: можно воздействовать на биологию (фармакотерапия, физическая активность, питание, сон), и это повлияет на психологическое состояние; можно воздействовать на психологию (психотерапия, изменение когнитивных паттернов, эмоциональная регуляция), и это повлияет на биологию. Оба направления воздействия легитимны и эффективны именно потому, что психологическое и биологическое, будучи различными уровнями, остаются аспектами единой системы и взаимно определяют друг друга через многочисленные петли обратной связи.
Философская традиция размышления об отношении ума и тела, сознания и материи насчитывает тысячелетия, и современная наука о стрессе вносит свой вклад в эту дискуссию, предоставляя эмпирические данные о конкретных механизмах связи психологического и биологического. Декартовский дуализм, постулировавший принципиальное различие между мыслящей субстанцией и протяжённой субстанцией и оставлявший проблему их взаимодействия неразрешённой, уступил место более нюансированным концепциям, признающим психофизическое единство человека. Феноменологическая традиция, идущая от Эдмунда Гуссерля и Мориса Мерло-Понти, подчёркивала «воплощённость» сознания, его неотделимость от телесного бытия. Современная когнитивная нейронаука развивает эту интуицию, демонстрируя, как телесные состояния формируют психические процессы: интероцепция — восприятие сигналов от внутренних органов — влияет на эмоциональный опыт; телесная поза влияет на когнитивные оценки; изменение паттерна дыхания меняет эмоциональное состояние. Эти данные поддерживают концепцию воплощённого познания, согласно которой разум не просто «помещён» в тело, но конституирован телесностью, и психологические процессы неразрывно связаны с соматическими.
Для понимания стресса особенно важен тот аспект эмерджентности, который касается возможности нисходящей причинности — влияния высших уровней организации на низшие. Редукционистская картина предполагает только восходящую причинность: молекулы определяют свойства клеток, клетки определяют работу органов, органы определяют функции организма, и так далее. Однако эмерджентные системы допускают и обратное направление влияния: паттерны организации на высшем уровне могут модулировать процессы на низшем. В контексте стресса нисходящая причинность проявляется в феномене психосоматических влияний: мысли и эмоции — психологические феномены — вызывают изменения в биологии организма. Когнитивная переоценка угрожающей ситуации как безопасной приводит к снижению активности миндалины и уровня кортизола. Хроническое переживание одиночества изменяет экспрессию генов в иммунных клетках. Психотерапия, изменяющая паттерны мышления и эмоциональной регуляции, нормализует нейронную активность и воспалительные маркёры. Эти факты демонстрируют, что психологический уровень не является эпифеноменом — бессильной тенью биологических процессов, — но обладает реальной каузальной силой, способностью влиять на нижележащие уровни.
Признание эмерджентности имеет важные следствия для понимания индивидуальных различий в переживании стресса. Если бы стресс был «просто биологией», то одинаковые биологические параметры — скажем, одинаковый уровень кортизола — предполагали бы одинаковое субъективное переживание. Однако это очевидно не так: два человека с одинаковыми биологическими маркёрами могут переживать стресс совершенно по-разному в зависимости от того, как они интерпретируют свою ситуацию, какой смысл ей придают, какие ресурсы для совладания у них имеются. Субъективное переживание зависит не только от «сырых» биологических данных, но и от того, как эти данные обрабатываются когнитивной системой, какое место они занимают в картине мира данного человека, в его нарративе о себе и своей жизни. Эта интерпретативная, смыслопорождающая функция является характерной особенностью психологического уровня и не может быть редуцирована к нейрофизиологии, хотя и реализуется нейронными механизмами. Понимание этого позволяет объяснить, почему «объективно» одинаковые стрессоры вызывают столь различные реакции у разных людей и почему психологические интервенции, изменяющие интерпретацию, а не биологию непосредственно, могут быть столь эффективны.
Концепция эмерджентности также помогает понять терапевтический потенциал работы на психологическом уровне и объясняет, почему «просто разговор» может приводить к реальным изменениям в теле. Психотерапия часто воспринимается как нечто «нематериальное» в противопоставлении «материальным» фармакологическим вмешательствам. Однако с позиции эмерджентности это противопоставление ложно: психотерапия работает с эмерджентными психологическими паттернами — убеждениями, схемами интерпретации, стратегиями совладания, — и изменение этих паттернов транслируется в изменения на биологическом уровне через механизмы нисходящей причинности. Нейровизуализационные исследования демонстрируют, что успешная психотерапия изменяет паттерны активации мозга, нормализует функциональную связность между областями, участвующими в эмоциональной регуляции, модулирует нейрохимические системы. Эти изменения не менее «материальны», чем изменения, вызванные препаратами; просто точка входа в систему находится на другом уровне. Признание реальности и каузальной эффективности психологического уровня обосновывает легитимность психологических интервенций и объясняет их биологические эффекты.
Философская основа, заложенная концепцией эмерджентности, будет сопровождать все последующие разделы курса, касающиеся психологических аспектов стресса. Когда мы обсуждаем когнитивную оценку стрессоров, эмоциональные реакции, стратегии совладания, мы работаем с эмерджентными феноменами, которые несводимы к своей биологической основе, хотя и неотделимы от неё. Понимание этой несводимости защищает от наивного редукционизма, который обесценивает субъективный опыт и психологические вмешательства. Понимание неотделимости от биологии защищает от дуалистического игнорирования телесного измерения стресса. Интегративная позиция, признающая многоуровневую природу стресса с эмерджентными свойствами на каждом уровне, обеспечивает наиболее адекватную концептуальную рамку как для исследования, так и для практической помощи людям, переживающим стресс.
4.2 Восприятие и когнитивная оценка стрессора
Традиционное представление о стрессе как о реакции организма на внешнее воздействие содержит в себе неявное допущение, что определённые события являются стрессорами объективно, независимо от того, кто их переживает. Публичное выступление, экзамен, конфликт с начальником — эти ситуации часто рассматриваются как «стрессовые по определению», порождающие стресс-реакцию у любого человека, который с ними сталкивается. Однако повседневный опыт и систематические исследования опровергают эту интуицию: одна и та же ситуация может вызывать интенсивный стресс у одного человека, умеренное возбуждение у другого и полное равнодушие у третьего. Более того, один и тот же человек может реагировать на одну и ту же ситуацию по-разному в зависимости от контекста, своего текущего состояния, доступных ресурсов. Эти наблюдения привели к фундаментальному переосмыслению природы стресса, кульминацией которого стала транзактная модель стресса, разработанная Ричардом Лазарусом и Сьюзан Фолкман в восьмидесятых годах двадцатого века. Согласно этой модели, стресс не локализован ни в ситуации, ни в человеке, но возникает в транзакции между ними — в процессе взаимодействия человека с окружающей средой, центральным элементом которого является когнитивная оценка.
Когнитивная оценка, согласно модели Лазаруса, представляет собой процесс, посредством которого человек определяет значение события для своего благополучия и свои возможности справиться с ситуацией. Эта оценка не является чисто интеллектуальным, рациональным анализом, проводимым хладнокровно и отстранённо; она часто происходит быстро, автоматически, с минимальным участием сознательного размышления, и она неразрывно связана с эмоциональным откликом. Лазарус выделял два основных компонента оценки, которые он назвал первичной и вторичной оценкой, хотя эти термины не указывают на временну́ю последовательность — оба процесса могут происходить практически одновременно и взаимно влиять друг на друга. Первичная оценка касается значения события: представляет ли оно угрозу, утрату или вызов? Угроза подразумевает антиципацию возможного вреда, который ещё не наступил, но может наступить; утрата означает, что вред уже произошёл; вызов предполагает ситуацию, требующую мобилизации ресурсов, но не воспринимаемую как превышающую возможности человека. Эти различные оценки порождают различные эмоциональные и физиологические паттерны: угроза ассоциирована со страхом и тревогой, утрата — с грустью и горем, вызов — с возбуждением и предвкушением.
Вторичная оценка касается ресурсов и возможностей совладания: что человек может сделать в данной ситуации, какие стратегии доступны, насколько вероятен успешный исход. Именно соотношение между воспринимаемыми требованиями ситуации (первичная оценка) и воспринимаемыми ресурсами для совладания (вторичная оценка) определяет, будет ли ситуация переживаться как стрессовая. Если воспринимаемые требования превышают воспринимаемые ресурсы, возникает переживание стресса; если ресурсы воспринимаются как достаточные или превышающие требования, ситуация может восприниматься как вызов, возможность, даже удовольствие. Примечательно, что речь идёт именно о воспринимаемых требованиях и ресурсах, а не об объективных: человек может обладать всеми необходимыми навыками для успешного выполнения задачи, но если он не верит в свою компетентность, он будет переживать стресс. И наоборот, человек с несколько завышенной самооценкой может спокойно подходить к ситуациям, которые объективно превышают его возможности. Эта субъективность оценки имеет как адаптивные, так и дезадаптивные последствия.
Процесс когнитивной оценки разворачивается чрезвычайно быстро, часто в течение долей секунды, и значительная его часть протекает за пределами сознательного контроля. Нейробиологические исследования демонстрируют, что миндалина — ключевая структура мозга, участвующая в обработке эмоционально значимой информации и особенно угроз, — способна оценивать стимулы и запускать физиологическую реакцию ещё до того, как информация достигает коры и становится доступной для сознательного анализа. Существует быстрый подкорковый путь обработки угрожающей информации, идущий от сенсорных органов через таламус напрямую к миндалине, минуя более медленную и детальную корковую обработку. Этот механизм имеет очевидную эволюционную логику: в ситуации реальной опасности — встречи с хищником, падения, приближения угрожающего объекта — миллисекунды могут определять разницу между жизнью и смертью, и организм не может позволить себе роскошь неторопливого сознательного анализа. Лучше ложноположительная реакция — испугаться того, что оказалось безобидным, — чем ложноотрицательная — не среагировать на реальную угрозу. Однако в современной жизни, где большинство стрессоров не являются непосредственными физическими угрозами, этот механизм может порождать избыточные стресс-реакции на ситуации, которые при сознательном анализе оказались бы не столь угрожающими.
Сознательная, рефлексивная оценка подключается после первичной автоматической реакции и может модулировать, корректировать или переоценивать исходную интерпретацию. Этот процесс переоценки является ключевым механизмом эмоциональной регуляции и мишенью многих психотерапевтических вмешательств. Классический пример: человек просыпается ночью от неожиданного звука. Мгновенная автоматическая оценка интерпретирует звук как потенциальную угрозу — возможно, кто-то проник в дом; симпатическая нервная система активируется, сердце начинает биться быстрее, мышцы напрягаются, внимание обостряется. Затем включается сознательный анализ: откуда звук, что он напоминает, есть ли другие признаки опасности. Человек вспоминает, что забыл закрыть форточку, и порыв ветра, вероятно, что-то уронил в соседней комнате; или он узнаёт характерные звуки кота, сбившего что-то со стола. Угроза переоценивается как ложная тревога, и физиологическое состояние начинает нормализоваться — хотя и не мгновенно, поскольку гормоны стресса уже выброшены в кровь и требуется время для их метаболизма. Этот пример иллюстрирует взаимодействие быстрой автоматической и медленной сознательной оценки в формировании и модуляции стресс-ответа.
Модель Лазаруса признаёт, что когнитивная оценка не является одномоментным событием, но представляет собой непрерывный процесс, разворачивающийся во времени по мере развития ситуации и изменения доступной информации. Первоначальная оценка может пересматриваться многократно на основе новых данных, обратной связи от собственных действий, изменений в контексте. Лазарус использовал термин «переоценка» для обозначения этих последующих модификаций первоначальной оценки. Переоценка может происходить спонтанно, по мере того как разворачивается ситуация и появляется новая информация, или намеренно, как сознательная стратегия эмоциональной регуляции. Когнитивная психотерапия во многом основана на идее, что люди могут научиться осознавать свои автоматические оценки, подвергать их критическому анализу и формировать более адаптивные интерпретации. Способность к гибкой переоценке является важным аспектом психологической устойчивости: люди, способные пересматривать свои первоначальные катастрофические интерпретации, обнаруживать альтернативные объяснения, находить возможности даже в трудных ситуациях, демонстрируют лучшие показатели психического и физического здоровья.
Индивидуальные различия в паттернах когнитивной оценки формируются на протяжении жизни под влиянием темперамента, раннего опыта, культурного контекста, предшествующих стрессов и успехов совладания. Дети, выросшие в небезопасной среде, могут развить паттерн гипербдительности — склонность интерпретировать неоднозначные сигналы как угрожающие, — который был адаптивен в их изначальном контексте, но становится источником избыточного стресса в более безопасных обстоятельствах взрослой жизни. Травматический опыт может формировать устойчивые схемы интерпретации, в которых мир воспринимается как опасный, другие люди как потенциально враждебные, а собственные возможности совладания как недостаточные. Культурные факторы также влияют на оценку: в одних культурах определённые ситуации — например, публичная критика — воспринимаются как крайне угрожающие для социального «лица», тогда как в других они рассматриваются более спокойно. Эти индивидуальные и культурные различия объясняют вариабельность стресс-реакций и подчёркивают, что понимание когнитивной оценки конкретного человека необходимо для понимания его переживания стресса.
Концепция когнитивной оценки имеет важные терапевтические импликации, которые реализуются в различных подходах когнитивной и когнитивно-поведенческой психотерапии. Если стресс определяется не столько самой ситуацией, сколько её интерпретацией, то изменение интерпретации может изменить переживание стресса без изменения объективных обстоятельств. Работа с автоматическими мыслями — мгновенными интерпретациями, возникающими в ответ на ситуацию, — является центральным элементом когнитивной терапии Аарона Бека. Пациент учится осознавать свои автоматические оценки, проверять их на соответствие реальности, формулировать альтернативные, более адаптивные интерпретации. Рационально-эмоциональная терапия Альберта Эллиса фокусируется на иррациональных убеждениях — глубинных предположениях о том, какими должны быть мир, другие люди и сам человек, — которые делают определённые ситуации катастрофическими. Терапия принятия и ответственности предлагает иной подход: вместо изменения содержания мыслей развивается способность относиться к мыслям как к мыслям — психологическим событиям, которые не обязательно отражают реальность и не требуют немедленного реагирования. Все эти подходы, при их различиях, объединены признанием центральной роли когнитивной оценки в генерации и поддержании стресса.
Связь когнитивной оценки с биологическими процессами стресс-ответа представляет собой яркий пример нисходящей причинности, обсуждавшейся в контексте эмерджентности. Когда человек оценивает ситуацию как угрожающую, эта психологическая оценка транслируется в активацию нейронных цепей, связывающих префронтальную кору с миндалиной и гипоталамусом, что запускает каскад нейроэндокринных событий: выброс кортикотропин-рилизинг-гормона, активацию симпатической нервной системы, продукцию кортизола. Тот же самый физиологический стимул — например, учащённое сердцебиение — может быть интерпретирован по-разному в зависимости от контекста и запускать различные паттерны активации. Человек на беговой дорожке интерпретирует тахикардию как естественный результат физической нагрузки; тот же человек, внезапно ощутивший учащение пульса в спокойной ситуации, может интерпретировать его как признак надвигающегося сердечного приступа или панической атаки, что усиливает тревогу и дополнительно ускоряет ритм — классический порочный круг паники. Исследования показывают, что инструкция интерпретировать физиологическое возбуждение как вызов, а не угрозу, меняет не только субъективные отчёты, но и биологические параметры — профиль сердечно-сосудистой реакции, уровни кортизола, маркёры воспаления.
Концепция когнитивной оценки будет детально развита в последующих модулях курса, посвящённых психологии стресса, однако её введение уже на этом этапе необходимо для понимания того, как биологические и психологические уровни анализа интегрируются в единую картину. Стресс не является чисто биологическим феноменом, механически запускаемым внешними стимулами; он не является и чисто психологическим, существующим только в субъективном переживании вне связи с телом. Стресс возникает на пересечении биологического и психологического, в транзакции между организмом и средой, опосредованной когнитивной оценкой. Эта интегративная перспектива имеет фундаментальное значение как для понимания механизмов стресса, так и для разработки эффективных стратегий его преодоления, которые должны адресоваться обоим уровням — и биологическому, и психологическому.
4.3 Эмоциональные реакции — сигналы и регуляторы
Эмоции, сопровождающие переживание стресса, — страх, тревога, гнев, грусть, отчаяние, беспомощность — часто воспринимаются как неприятные помехи, от которых хотелось бы избавиться. Культурные установки, особенно в западных обществах с их акцентом на рациональность и контроль, нередко представляют эмоции как нечто низшее по сравнению с разумом, как атавизм животной природы, препятствующий эффективному функционированию. С этой точки зрения идеальным ответом на стресс был бы хладнокровный, бесстрастный анализ ситуации и рациональное принятие решений, не затуманенное эмоциональными переживаниями. Однако современная аффективная наука — междисциплинарная область, изучающая эмоции с позиций психологии, нейронауки, эволюционной биологии, — предлагает принципиально иной взгляд: эмоции не являются помехой адаптации, но представляют собой эволюционно сформированную систему быстрой оценки и реагирования, критически важную для выживания и благополучия. Эмоции при стрессе — это не шум, а сигнал; не препятствие, а ресурс. Понимание информационной и регуляторной функции эмоций фундаментально изменяет подход к управлению стрессом: вместо подавления эмоций речь идёт об их понимании, принятии и конструктивном использовании.
Эмоции можно понимать как интегрированные паттерны ответа, включающие субъективное переживание, физиологические изменения, когнитивные сдвиги и поведенческие тенденции, которые мобилизуют организм для определённого типа действия в ответ на значимые события. Страх, например, представляет собой не просто неприятное чувство, но комплексный ответ, включающий специфическое переживание ощущения угрозы, физиологическую активацию симпатической нервной системы с характерными соматическими проявлениями, сужение внимания на источнике опасности и готовность к бегству или замиранию. Гнев включает переживание нарушения границ или несправедливости, физиологическую мобилизацию иного профиля с характерным повышением кровенаполнения периферии, когнитивный фокус на источнике фрустрации и готовность к конфронтации или агрессии. Грусть сопровождается переживанием утраты, физиологическим «замедлением» с преобладанием парасимпатических влияний, когнитивным обращением к утраченному и поведенческим отступлением от активности. Каждая базовая эмоция представляет собой эволюционно отточенный «пакет» ответов, подготавливающий организм к определённому типу ситуаций.
Информационная функция эмоций состоит в том, что они сообщают человеку о значении происходящего для его благополучия быстрее и часто точнее, чем сознательный анализ. Эмоция — это сигнал: страх говорит «здесь опасность, нужно защититься»; гнев сообщает «граница нарушена, нужно восстановить справедливость»; грусть указывает «произошла потеря, нужно время для горевания и восстановления»; отвращение предупреждает «это может быть вредно, избегай»; радость подтверждает «это хорошо, продолжай»; интерес направляет «это стоит исследовать». Эта сигнальная функция особенно ценна в условиях неопределённости и дефицита времени, когда сознательный анализ всех факторов невозможен. Интуитивное «чувствование» ситуации, которое люди описывают как «внутренний голос» или «шестое чувство», часто представляет собой результат неосознаваемой эмоциональной оценки, интегрирующей тонкие сигналы, которые не достигли порога сознательного восприятия. Исследования в области принятия решений демонстрируют, что люди с нарушениями эмоциональной обработки вследствие повреждения определённых областей мозга принимают худшие решения, несмотря на сохранный интеллект, — именно потому, что они лишены эмоциональных «маркёров», направляющих выбор.
Связь эмоций с биологическими процессами стресс-ответа является двунаправленной: эмоции не только сопровождают физиологические изменения, но и активно регулируют их. Переживание страха запускает симпатоадреналовую активацию — выброс адреналина, учащение сердцебиения, мобилизацию мышц. Хроническое переживание тревоги поддерживает повышенный уровень кортизола и провоспалительных цитокинов. Грусть и горе ассоциированы с характерными изменениями иммунной функции и повышенным риском соматических заболеваний. Однако и позитивные эмоции оказывают биологические эффекты: переживание радости, интереса, благодарности, любви ассоциировано с иным профилем вегетативной активации, иным гормональным паттерном, иными иммунными характеристиками. Исследования показывают, что позитивные эмоции способствуют более быстрому восстановлению после стресса, нормализации сердечно-сосудистых параметров, улучшению иммунной функции. Барбара Фредриксон предложила «расширяющую и созидающую теорию» позитивных эмоций, согласно которой они расширяют когнитивный и поведенческий репертуар, способствуют накоплению ресурсов и противодействуют сужающим эффектам негативных эмоций.
Подавление эмоций, нередко рассматриваемое как проявление самоконтроля и силы воли, в действительности оказывается неэффективной и потенциально вредной стратегией управления эмоциональным опытом. Исследования Джеймса Гросса и коллег, изучавших различные стратегии эмоциональной регуляции, демонстрируют, что подавление — сознательное усилие не выражать эмоцию и не думать о ней — требует значительных когнитивных ресурсов, истощает саморегуляторный потенциал и, парадоксально, может усиливать физиологическую реакцию, несмотря на уменьшение внешних проявлений. Человек, подавляющий страх, выглядит спокойнее, но его симпатическая активация может быть даже выше, чем у человека, открыто переживающего страх. Более того, подавление нарушает когнитивную обработку и память, ухудшает социальное функционирование, поскольку партнёры по общению ощущают дискомфорт от несоответствия между эмоциональным состоянием человека и его поведением. Хроническое подавление эмоций ассоциировано с рядом негативных последствий для здоровья, включая повышенный риск сердечно-сосудистых заболеваний, нарушения иммунной функции, депрессивные симптомы. Тело продолжает реагировать на эмоцию, даже когда сознание отказывается её признавать.
Альтернативой подавлению является эмоциональная регуляция — более широкое понятие, обозначающее процессы, посредством которых люди влияют на то, какие эмоции они испытывают, когда их испытывают и как их выражают. В отличие от подавления, которое направлено на устранение эмоции после её возникновения, эффективная эмоциональная регуляция может осуществляться на разных этапах эмоционального процесса. Выбор ситуации означает активное приближение к ситуациям, вызывающим желательные эмоции, и избегание ситуаций, вызывающих нежелательные. Модификация ситуации предполагает изменение объективных характеристик ситуации для изменения её эмоционального воздействия. Распределение внимания включает такие стратегии, как отвлечение или концентрация на определённых аспектах ситуации. Когнитивная переоценка, уже рассмотренная ранее, представляет собой изменение интерпретации ситуации. Наконец, модуляция ответа — это влияние на уже возникшую эмоциональную реакцию, которое может быть более или менее адаптивным. Исследования показывают, что когнитивная переоценка, осуществляемая на ранних этапах эмоционального процесса, является более эффективной и менее затратной стратегией, чем подавление, осуществляемое после полного развёртывания эмоции.
Концепция эмоционального интеллекта, популяризированная Дэниелом Гоулманом, но имеющая более глубокие академические корни в работах Питера Саловея и Джона Мэйера, подчёркивает значимость способности понимать и регулировать эмоции для успешного функционирования во всех сферах жизни. Эмоциональный интеллект включает несколько компонентов: способность воспринимать эмоции — распознавать их в себе и других по внешним признакам, речи, поведению; способность использовать эмоции для облегчения мышления — привлекать эмоции для направления внимания, принятия решений, творческого процесса; способность понимать эмоции — знать их причины, последствия, взаимосвязи, развитие во времени; способность управлять эмоциями — регулировать их интенсивность и выражение в соответствии с целями и контекстом. Люди с высоким эмоциональным интеллектом демонстрируют лучшую адаптацию к стрессу, более эффективное совладание, более устойчивые социальные отношения, более высокие показатели психического и физического здоровья. Эмоциональный интеллект, что важно, поддаётся развитию через обучение и практику, что открывает перспективы превентивных и терапевтических вмешательств.
Способность к осознаванию и называнию эмоций — тому, что психологи называют эмоциональной гранулярностью или эмоциональной дифференциацией, — представляет собой важный аспект эмоциональной компетентности, связанный с лучшими исходами при стрессе. Люди различаются по тому, насколько дифференцированно они воспринимают и описывают свои эмоциональные состояния: одни ограничиваются грубыми категориями «хорошо» и «плохо», другие способны различать множество оттенков — тревогу, беспокойство, страх, ужас, панику, или раздражение, недовольство, гнев, ярость, бешенство. Исследования показывают, что люди с высокой эмоциональной гранулярностью лучше регулируют свои эмоции, реже прибегают к дезадаптивным стратегиям вроде злоупотребления алкоголем или агрессии, демонстрируют меньшую реактивность на стресс. Возможное объяснение состоит в том, что точная идентификация эмоции позволяет выбрать более адекватную стратегию совладания: ответ на тревогу отличается от ответа на грусть или гнев, и смешение этих состояний в недифференцированное «плохо» затрудняет выбор эффективного действия. Практики осознанности и психотерапия развивают эмоциональную гранулярность через обращение внимания к тонким различиям эмоциональных состояний.
Алекситимия — клинический конструкт, обозначающий затруднение в идентификации и описании собственных эмоциональных состояний, — представляет собой противоположный полюс эмоциональной компетентности и ассоциирована с рядом неблагоприятных последствий для здоровья. Люди с алекситимией испытывают эмоции, но не могут их ясно осознать и вербализовать; вместо эмоциональных переживаний они нередко отмечают соматические симптомы — напряжение, боль, дискомфорт, — не связывая их с эмоциональными причинами. Алекситимия ассоциирована с соматоформными расстройствами, при которых психологический дистресс выражается через телесные симптомы; с зависимостями, где вещества используются для регуляции неопознанных эмоциональных состояний; с рядом соматических заболеваний, включая сердечно-сосудистые и желудочно-кишечные. Связь алекситимии с хроническим стрессом и его последствиями подчёркивает важность эмоциональной осведомлённости для здоровья: неспособность осознать эмоцию не устраняет её физиологических эффектов, но лишает человека возможности адекватно на неё ответить. Терапевтическая работа с алекситимией направлена на развитие способности замечать, идентифицировать и называть эмоциональные состояния.
Интеграция эмоциональной осведомлённости и регуляции в целостный подход к управлению стрессом представляет собой одну из ключевых задач психологических интервенций. Эмоции при стрессе — не враги, которых нужно побеждать, и не слабости, которых нужно стыдиться, но союзники, предоставляющие важную информацию и мобилизующие ресурсы. Страх сигнализирует об угрозе и готовит к защите; гнев указывает на нарушение границ и мобилизует для их восстановления; грусть говорит о потере и создаёт условия для горевания и перестройки; тревога предупреждает о неопределённости и побуждает к подготовке. Принятие этих эмоций, вместо борьбы с ними, парадоксальным образом часто способствует их регуляции: сопротивление эмоции требует ресурсов и создаёт дополнительное напряжение, тогда как принятие позволяет эмоции естественно развернуться и завершиться. Одновременно развитие навыков регуляции — переоценки, распределения внимания, модуляции физиологического возбуждения через дыхание и релаксацию — расширяет репертуар возможных ответов и предотвращает застревание в неконструктивных эмоциональных состояниях. Этот интегративный подход, сочетающий принятие эмоций с их умелой регуляцией, развивается в различных терапевтических традициях и представляет собой современный стандарт работы с эмоциональными аспектами стресса.
4.4 Когнитивные процессы под давлением стресса
Когнитивные функции — внимание, память, мышление, принятие решений — не остаются неизменными под воздействием стресса, но претерпевают характерные трансформации, которые имеют как адаптивное значение в контексте острой угрозы, так и потенциально дезадаптивные последствия при хроническом стрессе. Традиционное противопоставление эмоций и разума, представляющее их как антагонистические силы, где эмоции «затуманивают» ясное мышление, является существенным упрощением. В действительности эмоции и когнитивные процессы глубоко интегрированы на нейронном уровне: лимбические структуры, ответственные за эмоциональную обработку, анатомически и функционально связаны с префронтальной корой, обеспечивающей высшие когнитивные функции, и эти связи модулируют работу друг друга в обоих направлениях. Стресс изменяет баланс между этими системами, сдвигая его в сторону более быстрых, автоматических, эмоционально-управляемых процессов и ослабляя влияние более медленных, контролируемых, аналитических. Этот сдвиг не является дисфункцией — он представляет собой адаптивную перенастройку когнитивной системы на режим быстрого реагирования, оптимизированный для выживания в условиях непосредственной угрозы. Однако в современной жизни, где большинство стрессоров требуют сложного анализа и стратегического планирования, а не мгновенного действия, этот эволюционно древний паттерн может становиться источником проблем.
Внимание — базовый когнитивный процесс, определяющий, какая информация из потока сенсорных данных будет отобрана для дальнейшей обработки, — демонстрирует характерные изменения при стрессе, которые могут быть описаны метафорой сужения или туннелирования. В состоянии острого стресса внимание концентрируется на источнике угрозы и непосредственно релевантной информации, тогда как периферические, менее значимые в данный момент стимулы отфильтровываются. Этот феномен, известный как «туннельное зрение» или «оружейный фокус» в контексте свидетельских показаний о преступлениях, имеет очевидную адаптивную логику: когда на вас несётся хищник, важно отслеживать его траекторию, а не любоваться окружающим пейзажем. Исследования демонстрируют, что эмоционально значимые стимулы, особенно угрожающие, захватывают внимание приоритетно, даже когда человек пытается игнорировать их и сосредоточиться на другой задаче. Нейробиологические механизмы этого эффекта включают модуляцию сенсорных областей коры миндалиной, которая усиливает обработку эмоционально релевантной информации. Однако обратной стороной этой концентрации является утрата периферического внимания: человек в состоянии стресса может упускать информацию, которая объективно важна, но не связана непосредственно с воспринимаемой угрозой, что может приводить к ошибкам суждения и действия.
Память — способность кодировать, хранить и извлекать информацию — также демонстрирует характерный паттерн изменений при стрессе, который нельзя описать однозначно как улучшение или ухудшение, но который отражает специфическую перенастройку системы. Острый стресс усиливает консолидацию — процесс переноса информации из кратковременной памяти в долговременную — для эмоционально значимого материала. Это объясняет, почему травматические события запоминаются ярко и детально, иногда с почти фотографической чёткостью, и могут сохраняться в памяти на протяжении всей жизни. Нейроэндокринные механизмы этого эффекта включают модуляцию гиппокампальной консолидации адреналином и кортизолом: эти гормоны, действуя через норадренергическую систему и глюкокортикоидные рецепторы в гиппокампе, усиливают молекулярные процессы, лежащие в основе формирования долговременных воспоминаний. Однако одновременно с усилением памяти на центральные, эмоционально значимые аспекты события наблюдается ослабление памяти на периферические детали. Человек, ставший жертвой ограбления, может с поразительной точностью помнить лицо преступника или оружие в его руке, но совершенно не помнить, во что он был одет или какие ещё люди находились рядом.
Хронический стресс, в отличие от острого, оказывает преимущественно негативное влияние на память, особенно на гиппокамп-зависимые формы памяти, включающие запоминание новой информации и пространственную ориентацию. Продолжительное воздействие повышенного кортизола токсично для гиппокампа — структуры, критически важной для формирования новых эксплицитных воспоминаний. Нейровизуализационные исследования демонстрируют уменьшение объёма гиппокампа у людей с хроническим стрессом, депрессией, посттравматическим стрессовым расстройством. На клеточном уровне хронический стресс подавляет нейрогенез в зубчатой извилине гиппокампа — одной из немногих областей взрослого мозга, где продолжается образование новых нейронов, — и вызывает ретракцию дендритов существующих нейронов, уменьшая число синаптических контактов. Эти структурные изменения коррелируют с функциональными нарушениями: люди в состоянии хронического стресса часто жалуются на забывчивость, трудности концентрации, ощущение «мозгового тумана». Примечательно, что многие из этих изменений обратимы при устранении стресса, что свидетельствует о пластичности мозга и возможности восстановления.
Принятие решений и исполнительные функции — способность планировать, гибко переключаться между задачами, подавлять импульсивные реакции, удерживать информацию в рабочей памяти — зависят преимущественно от префронтальной коры и демонстрируют особую уязвимость к стрессу. Префронтальная кора, эволюционно наиболее молодая часть мозга, обеспечивающая высшие когнитивные функции, требует точного баланса нейромодуляторов — дофамина и норадреналина — для оптимального функционирования. Умеренные уровни этих веществ поддерживают эффективную работу, но при сильном стрессе их концентрации возрастают до уровней, которые нарушают функцию префронтальных сетей. Нейробиолог Эми Арнстен описала этот феномен как «химическое отключение» префронтальной коры при остром стрессе. Когда префронтальный контроль ослабевает, поведение становится более управляемым подкорковыми структурами — миндалиной, полосатым телом, — что проявляется в повышенной импульсивности, преобладании привычных реакций над гибким, контекстуально адаптированным поведением, затруднениях в планировании и принятии сложных решений. В ситуации реальной физической опасности быстрое автоматическое действие предпочтительнее долгого размышления, однако в современных ситуациях стресса — рабочем конфликте, финансовых трудностях, семейных проблемах — снижение аналитических способностей может вести к неоптимальным решениям.
Закон Йеркса-Додсона, сформулированный более века назад, описывает связь между уровнем возбуждения и эффективностью деятельности в форме перевёрнутой буквы U или колоколообразной кривой. При низком уровне активации организм недостаточно мобилизован, внимание рассеяно, мотивация слаба, и продуктивность страдает. При умеренном уровне активации достигается оптимум: системы работают эффективно, внимание сфокусировано, ресурсы мобилизованы. При чрезмерно высоком уровне активации, характерном для сильного стресса, эффективность снова падает — теперь уже из-за перегрузки, дезорганизации, «химического отключения» высших функций. Этот закон имеет важные практические следствия: умеренный стресс может быть продуктивен, повышая мотивацию и концентрацию, тогда как сильный стресс почти всегда вреден для когнитивной деятельности. Положение оптимума на кривой зависит от сложности задачи: для простых, хорошо выученных задач оптимальный уровень активации выше, для сложных, требующих творческого мышления и гибкости, — ниже. Это объясняет, почему даже умеренный стресс может нарушать выполнение сложных когнитивных задач.
Хронический стресс оказывает особенно пагубное влияние на когнитивные функции, поскольку кумулятивное воздействие гормонов стресса приводит к структурным изменениям в мозге, затрагивающим ключевые когнитивные структуры. Помимо уже упомянутых изменений в гиппокампе, хронический стресс вызывает ретракцию дендритов и снижение синаптической плотности в префронтальной коре, что лежит в основе устойчивых нарушений исполнительных функций. Одновременно в миндалине наблюдается противоположный паттерн — рост дендритов и увеличение синаптической плотности, что может объяснять усиленную эмоциональную реактивность и гипербдительность при хроническом стрессе. Эти структурные перестройки отражают адаптивный сдвиг когнитивной системы в направлении приоритета обнаружения угроз над сложным анализом — адаптивный в хронически опасной среде, но дезадаптивный в обычных условиях. Когнитивные симптомы депрессии — трудности концентрации, нерешительность, руминативное мышление, негативный когнитивный сдвиг — отчасти объясняются этими структурно-функциональными изменениями в мозге, вызванными хроническим стрессом.
Руминация — повторяющееся, навязчивое возвращение мыслей к источнику стресса, прокручивание в уме негативных событий, самокритичных оценок, пессимистических прогнозов — представляет собой особый паттерн когнитивной активности при стрессе, который парадоксальным образом усугубляет дистресс вместо того, чтобы его разрешать. На первый взгляд руминация может казаться попыткой решения проблемы через обдумывание, однако исследования показывают принципиальное отличие руминации от конструктивного анализа: руминация циклична, не ведёт к новому пониманию или плану действий, сопровождается негативным аффектом и усиливает его. Нейровизуализационные исследования связывают руминацию с активностью сети пассивного режима работы мозга — системы областей, активных при отсутствии внешней задачи и участвующих в самореферентном мышлении, — и с нарушением способности переключаться с этой сети на сети, ответственные за внешнюю направленность внимания. Руминация является сильным предиктором развития депрессии и тревожных расстройств, а также их затяжного течения. Терапевтические интервенции, направленные на прерывание руминации — техники осознанности, поведенческая активация, когнитивная реструктуризация, — показывают эффективность в уменьшении депрессивных симптомов.
Понимание когнитивных эффектов стресса имеет прямые практические следствия для организации деятельности, образования, профессиональной подготовки и терапии. В условиях, где критически важны сложное мышление, творческие решения, гибкость, необходимо минимизировать уровень стресса или обеспечивать восстановление когнитивных ресурсов. Это особенно значимо для профессий, связанных с принятием ответственных решений в стрессовых условиях — медицинских работников, пилотов, диспетчеров, аварийно-спасательных служб: понимание того, как стресс влияет на когнитивные функции, позволяет разрабатывать протоколы, компенсирующие эти эффекты. В терапевтическом контексте признание когнитивных нарушений при хроническом стрессе и депрессии важно для реалистичных ожиданий и для выбора интервенций: когда префронтальные функции ослаблены, сложные когнитивные техники могут быть труднодоступны, и более уместны поведенческие вмешательства или техники, работающие с телесным и эмоциональным уровнем. По мере восстановления префронтальной функции становятся возможны более сложные когнитивные интервенции.
4.5 Поведенческие ответы — копинг и его многообразие
Поведение представляет собой видимую, наблюдаемую манифестацию стресс-ответа, в которой внутренние процессы — когнитивные оценки, эмоциональные реакции, физиологическая мобилизация — транслируются в действия во внешнем мире. Именно через поведение человек взаимодействует со стрессором, пытаясь устранить его, уменьшить его воздействие, адаптироваться к нему или избежать его. Концепция копинга, или совладания, занимает центральное место в психологии стресса и обозначает поведенческие и когнитивные усилия, предпринимаемые человеком для управления требованиями ситуации, которые оцениваются как превышающие его ресурсы. Копинг отличается от автоматических защитных реакций — таких как рефлекторное отдёргивание руки от горячего предмета — тем, что включает элемент усилия, мобилизации ресурсов, выбора среди возможных стратегий. Разнообразие стратегий копинга огромно и отражает сложность человеческого поведенческого репертуара: от активного решения проблемы до эмоциональной разрядки, от поиска социальной поддержки до ухода в фантазии, от юмора до употребления психоактивных веществ. Понимание этого разнообразия и факторов, определяющих выбор и эффективность различных стратегий, является ключевой задачей психологии стресса.
Классическая классификация Лазаруса и Фолкман выделяет два основных типа копинга: проблемно-фокусированный и эмоционально-фокусированный. Проблемно-фокусированный копинг направлен на изменение ситуации, устранение или уменьшение стрессора: это может быть активное решение проблемы, планирование, поиск информации, переговоры, конфронтация с источником стресса. Эмоционально-фокусированный копинг направлен на регуляцию эмоционального состояния, вызванного стрессором, без непосредственного воздействия на саму ситуацию: это может быть поиск эмоциональной поддержки, когнитивная переоценка, отвлечение, юмор, выражение эмоций, религиозная практика. Это разделение интуитивно понятно и соответствует двум фундаментальным задачам, которые человек решает при столкновении со стрессом: изменить ситуацию, если это возможно, и справиться с эмоциональными последствиями, если ситуация неизменяема или пока она не изменена. Однако современные исследования подчёркивают, что это разделение не является жёстким: многие стратегии совладания служат одновременно обеим целям, и эффективное совладание обычно включает элементы обоих типов.
Третье важное измерение копинга — различие между приближением и избеганием — описывает, направлена ли стратегия на конфронтацию со стрессором или на уход от него. Копинг приближения включает активное обращение к проблеме, сбор информации о ней, обдумывание возможных решений, эмоциональное выражение и проработку переживаний. Копинг избегания включает стратегии ухода от стрессора или мыслей о нём: отвлечение, отрицание, подавление мыслей и чувств, поведенческий уход из ситуации, употребление веществ для изменения состояния сознания. Долгое время в литературе преобладало представление о том, что копинг приближения адаптивен, тогда как копинг избегания дезадаптивен. Однако современные исследования показывают более нюансированную картину: краткосрочное избегание может быть адаптивным, когда ресурсы для активного совладания недостаточны или когда стрессор временный и нетрансформируемый; проблема возникает, когда избегание становится хроническим и единственным паттерном, блокирующим возможность адаптации. Гибкость — способность использовать различные стратегии в зависимости от контекста — оказывается более важным предиктором благополучия, чем преобладание какого-либо одного стиля.
Вопрос об эффективности различных стратегий копинга не имеет универсального ответа, поскольку эффективность определяется соответствием стратегии характеру стрессора, контексту, ресурсам человека, временно́й перспективе. Центральным принципом является соответствие между типом стрессора и типом копинга. Проблемно-фокусированный копинг наиболее эффективен в ситуациях, которые поддаются изменению: когда человек сталкивается с экзаменом, он может учиться больше; когда возникает конфликт с коллегой, можно инициировать разговор и искать решение. Эмоционально-фокусированный копинг более уместен в ситуациях, которые невозможно изменить или которые уже произошли: потеря близкого человека, неизлечимый диагноз, стихийное бедствие. Попытка решить нерешаемую проблему ведёт к фрустрации и истощению, тогда как принятие неизменяемого и фокус на эмоциональной адаптации позволяет сохранить ресурсы и функционирование. Модель соответствия предсказывает, что несоответствие между контролируемостью ситуации и стратегией копинга ведёт к худшим исходам, и эта предсказание подтверждается эмпирически.
Социальная поддержка занимает особое место среди стратегий совладания, поскольку она является одним из наиболее мощных и универсальных буферов против негативных последствий стресса. Социальная поддержка включает несколько компонентов: эмоциональная поддержка — выражение заботы, сочувствия, принятия; инструментальная поддержка — практическая помощь, ресурсы, услуги; информационная поддержка — советы, информация, руководство; оценочная поддержка — обратная связь, подтверждение самооценки. Эффект социальной поддержки на здоровье документирован множеством исследований: люди с более сильными социальными связями демонстрируют лучшие показатели психического и физического здоровья, более низкую смертность от различных причин, лучшее восстановление после болезней и травм. Механизмы этого эффекта включают как прямые — социальное взаимодействие активирует нейроэндокринные системы, снижающие стресс-реактивность, стимулирует иммунную функцию, — так и опосредованные — социальная сеть обеспечивает информацию, помощь, контроль над поведением, связанным со здоровьем. Примечательно, что не только получение, но и оказание поддержки благоприятно для здоровья, что подчёркивает реципрокность и взаимность как важные аспекты социальных связей.
Дезадаптивные стратегии копинга, при всей своей неэффективности или вредоносности в долгосрочной перспективе, обычно приносят кратковременное облегчение, что и объясняет их устойчивость в поведенческом репертуаре. Употребление алкоголя и других психоактивных веществ для снятия стресса является классическим примером: алкоголь действительно временно снижает тревогу и создаёт ощущение расслабления через воздействие на ГАМКергическую систему, однако после окончания действия вещества тревога часто возвращается с большей интенсивностью, формируется толерантность и зависимость, страдает здоровье и социальное функционирование. Переедание или, напротив, ограничение питания могут служить механизмами регуляции эмоций, однако ведут к нарушениям пищевого поведения и соматическим проблемам. Социальная изоляция может казаться облегчением от требований социального взаимодействия, но лишает человека социальной поддержки и усугубляет депрессию. Руминация, рассмотренная ранее, может ощущаться как попытка понять и решить проблему, но в действительности лишь усиливает дистресс. Общий паттерн состоит в том, что стратегии, приносящие немедленное облегчение ценой долгосрочных издержек, имеют тенденцию закрепляться и становиться привычными, формируя порочные круги, которые трудно прервать.
Концепция копинговой гибкости, или адаптивной гибкости совладания, подчёркивает, что эффективность совладания определяется не столько репертуаром доступных стратегий или предпочтением какого-либо одного стиля, сколько способностью гибко выбирать стратегию в соответствии с требованиями ситуации и корректировать свой подход на основе обратной связи. Человек с высокой копинговой гибкостью способен распознать, является ли ситуация контролируемой или нет, и соответственно применить проблемно-фокусированный или эмоционально-фокусированный копинг; он способен оценить, приносит ли выбранная стратегия желаемый результат, и переключиться на альтернативную, если нет; он способен использовать различные стратегии для различных аспектов сложной ситуации. Исследования показывают, что копинговая гибкость ассоциирована с лучшими показателями адаптации к стрессу и психического здоровья. Это имеет терапевтические импликации: вместо обучения какой-либо одной «правильной» стратегии более продуктивно расширять репертуар доступных стратегий и развивать метакогнитивные навыки их гибкого применения.
Культурные факторы оказывают существенное влияние на репертуар копинговых стратегий, их социальную приемлемость и эффективность. В индивидуалистических культурах, характерных для западных обществ, ценятся автономия, самодостаточность, активное решение проблем, прямое выражение эмоций; стратегии, акцентирующие индивидуальный контроль и личную агентность, рассматриваются как наиболее адаптивные. В коллективистских культурах, характерных для многих азиатских обществ, большее значение придаётся гармонии в отношениях, социальному соответствию, непрямой коммуникации; стратегии, включающие поиск поддержки семьи, принятие, эмоциональную сдержанность, могут быть более адаптивными. Религиозный копинг — обращение к религиозной вере, молитве, религиозному сообществу — имеет различное значение в различных культурных контекстах. Это культурное разнообразие предостерегает от универсализации западных моделей «эффективного» копинга и подчёркивает необходимость культурной чувствительности в исследованиях и интервенциях. Эффективная стратегия — та, которая работает в данном контексте для данного человека, а не та, которая соответствует абстрактному стандарту.
Развитие навыков копинга представляет собой важную задачу как профилактических, так и терапевтических программ, направленных на повышение устойчивости к стрессу. Обучение копингу включает несколько компонентов: расширение осведомлённости о доступных стратегиях, поскольку многие люди ограничены в своём репертуаре не потому, что не способны к иным стратегиям, а потому, что просто не знают о них; развитие конкретных навыков, таких как решение проблем, коммуникация, релаксация, когнитивная реструктуризация; формирование метакогнитивных навыков оценки ситуации и выбора стратегии; практика применения навыков в различных ситуациях для достижения автоматизма. Программы обучения навыкам совладания демонстрируют эффективность в различных популяциях и контекстах: у школьников и студентов, у людей с хроническими заболеваниями, у профессионалов в стрессогенных профессиях, у людей, переживающих жизненные кризисы. Интеграция обучения копингу в более широкие программы укрепления психического здоровья представляет собой перспективное направление профилактики.
4.6 Сознательное и бессознательное в стресс-ответе
Рассмотрение психологических аспектов стресса было бы неполным без обращения к фундаментальному различию между сознательными и бессознательными процессами в формировании стресс-ответа. Традиционное представление о человеке как о рациональном агенте, сознательно оценивающем ситуации и выбирающем оптимальные стратегии действия, является существенным упрощением реальной картины. Значительная, возможно бо́льшая, часть психических процессов, включая восприятие угроз, эмоциональные реакции, поведенческие тенденции, протекает автоматически, без участия сознательного внимания и волевого контроля. Эта автоматическая обработка является не дефектом или примитивизмом, но эволюционно выработанным механизмом, обеспечивающим быстрое реагирование на значимые стимулы без когнитивно затратного сознательного анализа. Однако автоматизм имеет и обратную сторону: бессознательные процессы не подлежат прямому волевому изменению, они могут порождать реакции, неадекватные актуальному контексту, и они часто основаны на прошлом опыте, который может быть нерелевантен настоящему. Понимание соотношения сознательного и бессознательного в стресс-ответе критически важно как для объяснения паттернов реагирования, так и для разработки эффективных терапевтических стратегий.
Нейробиологические исследования демонстрируют существование быстрого подкоркового пути обработки угрожающей информации, который обеспечивает реакцию на опасность до формирования сознательного восприятия. Классические эксперименты Джозефа Леду показали, что информация о потенциально угрожающих стимулах может достигать миндалины — ключевой структуры эмоционального мозга — через прямой путь от таламуса, минуя кору больших полушарий. Этот «нижний путь» передаёт грубую, неполную информацию о стимуле, достаточную для первичной оценки потенциальной угрозы, и он работает быстрее, чем «верхний путь» через корковые области, обеспечивающий детальный анализ и осознанное восприятие. Таким образом, миндалина способна запустить физиологическую стресс-реакцию — активацию симпатической нервной системы, выброс адреналина, мышечное напряжение — ещё до того, как человек осознает, что он напуган, и тем более до того, как он сможет рационально оценить, обоснован ли страх. Субъективное переживание страха возникает с задержкой, когда информация от миндалины и от телесных изменений интегрируется с корковой обработкой. Эта последовательность — сначала реакция, потом осознание — противоречит интуитивному представлению о том, что мы сначала осознаём опасность и потом пугаемся.
Имплицитная память — форма памяти, влияющая на поведение и переживания без сознательного припоминания, — играет ключевую роль в формировании автоматических стресс-реакций. В отличие от эксплицитной, или декларативной памяти, которая включает осознанные воспоминания о событиях и фактах, имплицитная память проявляется в навыках, привычках, условных рефлексах, эмоциональных реакциях, не требуя осознания источника этих влияний. Травматический опыт, особенно пережитый в раннем детстве, когда системы эксплицитной памяти ещё незрелы, может кодироваться преимущественно в имплицитной форме. Человек может не иметь сознательных воспоминаний о травме, но его тело и эмоциональная система «помнят»: определённые стимулы — запахи, звуки, телесные ощущения, паттерны отношений — автоматически активируют защитные реакции страха, гнева, замирания, без осознания того, почему возникает эта реакция и с чем она связана. Феномен «телесной памяти», хотя и не вполне корректный как научный термин, указывает на реальность имплицитного соматического кодирования эмоционального опыта. Терапевтическая работа с травмой часто направлена на установление связи между этими имплицитными реакциями и их историческим источником, что позволяет переработать травматический материал и снизить его автоматическое влияние на настоящее.
Концепция автоматических мыслей в когнитивной терапии Аарона Бека указывает на другой уровень бессознательной обработки — когнитивный. Автоматические мысли — это мгновенные, непроизвольные интерпретации событий, которые возникают на периферии сознания или полностью за его пределами, но определяют эмоциональные и поведенческие реакции. Человек может не осознавать мысль «он меня критикует, значит, я неудачник», но переживать стыд и желание избежать ситуации. При систематическом самонаблюдении эти мысли могут быть выведены в поле сознания и подвергнуты критическому анализу. За автоматическими мыслями стоят более глубокие когнитивные структуры — убеждения, схемы, — которые формируются в раннем опыте и организуют интерпретацию всех последующих событий. Схема недостойности, например, сформированная в контексте критикующего или отвергающего воспитания, будет систематически направлять интерпретацию в сторону обнаружения признаков собственной несостоятельности и критики со стороны других. Эти глубинные структуры ещё менее доступны осознанию и более устойчивы к изменению, однако их трансформация и является целью глубинной психотерапевтической работы.
Роль детского опыта в формировании автоматических паттернов стресс-ответа заслуживает особого рассмотрения, поскольку именно в ранние годы жизни закладываются базовые схемы регуляции стресса и отношений с другими людьми. Теория привязанности, разработанная Джоном Боулби и эмпирически обоснованная Мэри Эйнсворт и последующими исследователями, описывает, как качество ранних отношений с ухаживающими взрослыми формирует внутренние рабочие модели — имплицитные представления о себе, других и отношениях, — которые затем направляют поведение и переживания на протяжении всей жизни. Ребёнок, чей опекун был доступен, отзывчив и надёжен, формирует безопасную привязанность и внутреннюю модель, в которой он сам достоин любви, другие люди надёжны, а мир в целом безопасен. Ребёнок, чей опекун был непредсказуем, отвергающ или пугающ, формирует небезопасную привязанность и соответствующие модели — себя как недостойного, других как ненадёжных или опасных, мира как угрожающего. Эти ранние модели функционируют преимущественно имплицитно, вне сознательного доступа, и автоматически активируются в ситуациях, связанных с близостью, зависимостью, уязвимостью — то есть именно в тех ситуациях, которые часто являются стрессорами во взрослой жизни.
Сознательная регуляция стресс-ответа возможна, однако она имеет свои ограничения и требует определённых условий для эффективности. Префронтальная кора, обеспечивающая сознательный контроль, способна модулировать активность подкорковых структур, включая миндалину, через нисходящие проекции. Этот механизм лежит в основе произвольной эмоциональной регуляции: когда человек сознательно переоценивает ситуацию или направляет внимание определённым образом, он активирует префронтальные области, которые, в свою очередь, снижают активность миндалины и уменьшают стресс-реакцию. Однако эта регуляция требует когнитивных ресурсов, которые могут быть истощены стрессом, усталостью, когнитивной нагрузкой. Как обсуждалось ранее, сильный стресс «отключает» префронтальную кору, лишая человека возможности сознательного контроля именно тогда, когда он наиболее необходим. Кроме того, сознательная регуляция эффективна лишь в отношении тех процессов, которые доступны осознанию: невозможно волевым усилием подавить имплицитную реакцию, о которой не знаешь. Эти ограничения объясняют, почему одного понимания недостаточно для изменения и почему терапевтическая работа часто должна выходить за пределы чисто когнитивных интервенций.
Терапевтические подходы к работе с бессознательными аспектами стресс-ответа различаются по теоретической ориентации, но объединены признанием необходимости выхода за пределы чисто рационального анализа. Психодинамические подходы, наследующие традиции психоанализа, фокусируются на исследовании бессознательных конфликтов, защитных механизмов, переносе ранних паттернов отношений на терапевтические отношения. Осознавание ранее бессознательного материала рассматривается как путь к освобождению от его автоматического влияния. Телесно-ориентированные подходы, такие как соматическое переживание Питера Левина или сенсомоторная психотерапия Пэт Огден, работают непосредственно с телесными аспектами стресс-ответа, помогая клиентам осознать и трансформировать соматические паттерны, закодированные в имплицитной памяти. Терапия десенсибилизации и переработки движениями глаз, или ДПДГ, использует билатеральную стимуляцию для облегчения переработки травматического материала, механизмы чего до конца не ясны, но эффективность документирована эмпирически. Практики осознанности развивают способность наблюдать автоматические реакции с позиции осведомлённого свидетеля, что само по себе может модулировать их интенсивность и автоматизм.
Интеграция сознательного и бессознательного в понимании стресса ведёт к более полной и нюансированной картине, чем предлагают подходы, фокусирующиеся исключительно на одном уровне. Когнитивно-поведенческая терапия, изначально фокусировавшаяся на сознательных мыслях и убеждениях, всё больше интегрирует работу с имплицитными процессами, эмоциями, телесными ощущениями. Психодинамические подходы, традиционно фокусировавшиеся на бессознательном, включают когнитивные и поведенческие элементы. Концепция «множественных уровней обработки» признаёт, что изменение может происходить на разных уровнях — когнитивном, эмоциональном, соматическом, реляционном — и что эффективная терапия должна адресоваться всем релевантным уровням. Для одних клиентов первичной точкой входа будет когнитивная работа, для других — эмоциональная или телесная; но в конечном счёте устойчивое изменение требует интеграции. Понимание того, что стресс-ответ формируется на множественных уровнях — от молекулярных процессов до сознательных смыслов, от автоматических реакций до намеренных действий, — создаёт основу для по-настоящему интегративного подхода к помощи людям, страдающим от последствий стресса.
Практические следствия понимания соотношения сознательного и бессознательного в стресс-ответе затрагивают не только терапию, но и профилактику, образование, самопомощь. На уровне профилактики это означает внимание к формирующим условиям раннего развития, которые закладывают базовые паттерны регуляции стресса. Программы поддержки родительства, раннее вмешательство при неблагоприятных условиях развития, создание безопасной среды в образовательных учреждениях — всё это способствует формированию адаптивных автоматических паттернов, которые будут служить человеку на протяжении жизни. На уровне самопомощи это означает признание того, что изменение паттернов стресс-ответа требует времени и практики: автоматизмы, формировавшиеся годами, не могут быть отменены одним актом понимания или волевого решения. Регулярная практика — будь то релаксация, осознанность, физическая активность или иные техники — постепенно формирует новые автоматические паттерны, которые со временем начинают активироваться столь же легко, как старые. Терпение к себе, признание ограничений волевого контроля и доверие к постепенному процессу изменения являются важными компонентами устойчивой работы над улучшением своих реакций на стресс.
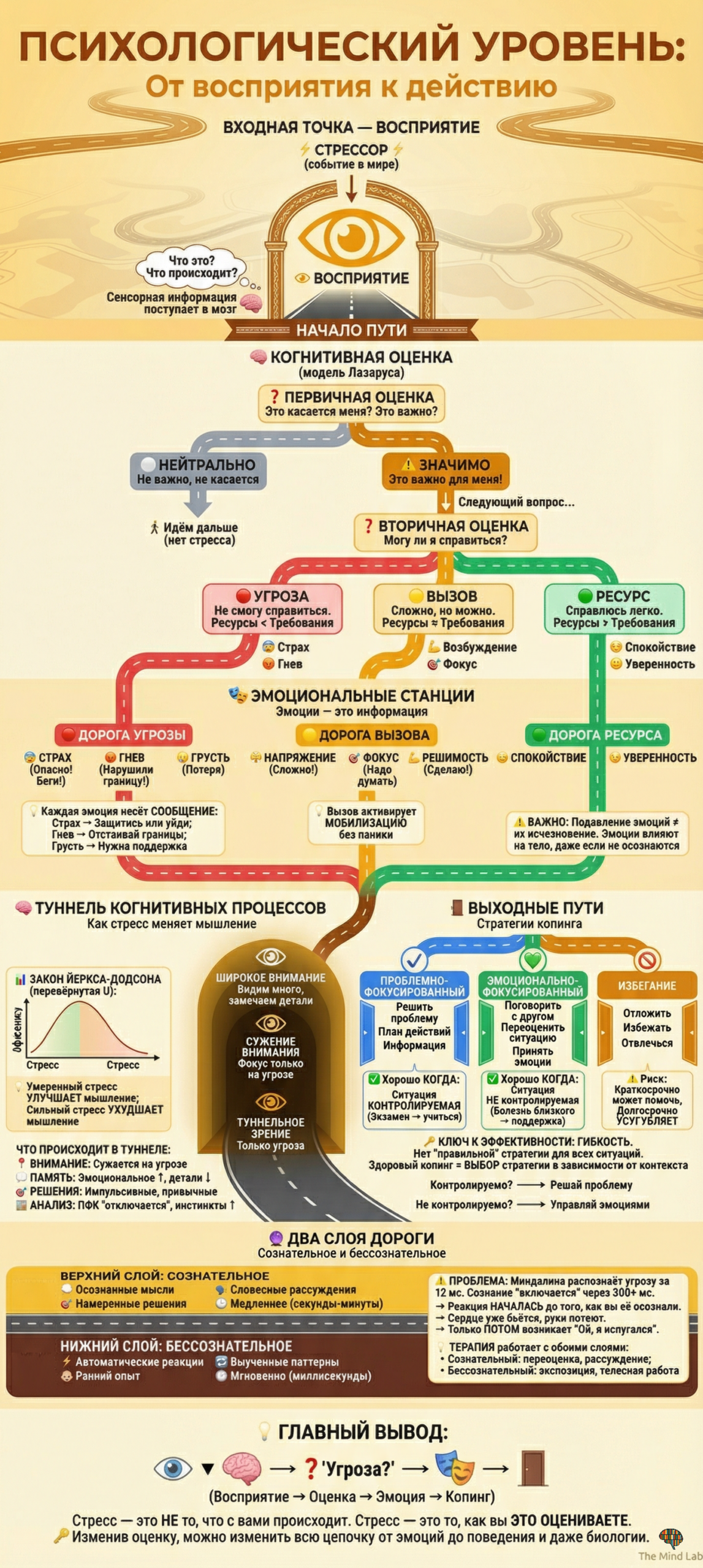
5. Социальный уровень — стресс в контексте отношений и общества
5.1 Межличностные отношения как источник и буфер стресса
Переход от психологического уровня анализа к социальному представляет собой ещё одно расширение перспективы, выводящее понимание стресса за пределы индивидуального организма в сферу отношений между людьми и социальных структур. Человек не является изолированным существом, переживающим стресс в одиночестве; он укоренён в сети социальных связей — семейных, дружеских, профессиональных, общественных, — которые фундаментальным образом формируют как его подверженность стрессорам, так и его способность справляться с ними. Парадокс социального измерения стресса состоит в том, что межличностные отношения одновременно являются наиболее мощным источником стресса и наиболее эффективным защитным фактором против его разрушительных последствий. Конфликты с близкими людьми, отвержение, утрата значимых отношений, одиночество — эти социальные стрессоры вызывают интенсивные физиологические реакции и ассоциированы с серьёзными последствиями для здоровья. Одновременно социальная поддержка — ощущение связи с другими, доступность помощи, принадлежность к сообществу — является одним из наиболее надёжных предикторов устойчивости к стрессу и хорошего здоровья. Понимание этой двойственности необходимо для полной картины стресса как биопсихосоциального феномена.
Эволюционная перспектива помогает понять, почему социальные стрессоры оказывают столь глубокое воздействие на человеческий организм. Homo sapiens эволюционировал как ультрасоциальный вид, выживание и репродуктивный успех которого критически зависели от принадлежности к группе. В условиях среды эволюционной адаптированности — африканской саванны плейстоцена — изгнание из группы было практически равнозначно смертному приговору: одинокий человек не мог эффективно защищаться от хищников, охотиться, добывать и сохранять пищу, заботиться о потомстве. Поэтому естественный отбор сформировал мозг, чрезвычайно чувствительный к социальным сигналам и воспринимающий угрозу социальным связям как экзистенциальную опасность. Нейробиологические исследования демонстрируют, что социальное отвержение активирует те же области мозга, которые реагируют на физическую боль, — в частности, переднюю поясную кору и переднюю островковую кору. Это не метафора: социальная боль нейробиологически родственна физической боли, что объясняет её интенсивность и почему люди готовы идти на значительные издержки, чтобы избежать остракизма. Современная социальная среда радикально отличается от среды эволюционной адаптированности, однако базовые нейробиологические механизмы остались прежними, и угроза социальной изоляции по-прежнему воспринимается организмом как угроза выживанию.
Социальная поддержка оказывает буферное действие на стресс-ответ, смягчая его интенсивность и сокращая продолжительность через множество физиологических механизмов. Присутствие поддерживающего другого человека — или даже его воображаемое присутствие — снижает реактивность гипоталамо-гипофизарно-надпочечниковой оси: в экспериментах люди, получающие социальную поддержку перед стрессовой задачей, демонстрируют меньший подъём кортизола, чем контрольные участники. Аналогичные эффекты наблюдаются в отношении сердечно-сосудистой реактивности: присутствие друга или партнёра снижает повышение артериального давления и частоты сердечных сокращений при стрессе. На уровне мозга социальная поддержка модулирует активность миндалины, снижая её реакцию на угрожающие стимулы. Воспалительные маркёры, повышающиеся при стрессе, также демонстрируют меньший подъём у людей с высоким уровнем социальной поддержки. Механизмы этих эффектов включают как прямые нейроэндокринные пути — выброс окситоцина при социальном контакте, который модулирует активность стресс-систем, — так и опосредованные когнитивные механизмы: поддержка других людей влияет на когнитивную оценку ситуации, делая её менее угрожающей, и на воспринимаемые ресурсы совладания.
Хроническое одиночество — субъективное переживание дефицита значимых социальных связей — представляет собой особенно опасный социальный стрессор с документированными последствиями для здоровья, сопоставимыми по масштабу с такими признанными факторами риска, как курение, ожирение, малоподвижный образ жизни. Эпидемиологические исследования показывают, что одинокие люди имеют повышенный риск сердечно-сосудистых заболеваний, инсульта, когнитивного снижения и деменции, депрессии и тревожных расстройств, а также преждевременной смерти от всех причин. Метаанализы оценивают, что социальная изоляция и одиночество повышают риск смертности примерно на двадцать шесть — тридцать процентов. Биологические механизмы этой связи включают хроническую активацию стресс-систем — повышенный кортизол, нарушение его циркадного ритма, симпатическую гиперактивацию, сниженную вариабельность сердечного ритма — и хроническое воспаление низкой интенсивности. Социальный нейробиолог Джон Качиоппо, посвятивший карьеру изучению одиночества, описывал его как состояние социального голода, при котором мозг переходит в режим гипербдительности, постоянно сканируя среду на предмет социальных угроз. Этот режим энергетически затратен и истощает адаптивные резервы, подобно тому как хронический стресс любой другой природы.
Качество социальных связей оказывается более значимым для здоровья и благополучия, чем их количество, что подчёркивает важность различения социальной изоляции и одиночества. Социальная изоляция — объективная характеристика, описывающая малое число социальных контактов и ограниченное участие в социальных активностях. Одиночество — субъективное переживание, которое может присутствовать даже при наличии многочисленных контактов, если они не удовлетворяют потребность в близости и принадлежности. Человек может быть социально изолирован, но не чувствовать себя одиноким, если имеющиеся немногие связи глубоки и значимы; и, напротив, может быть окружён людьми, но чувствовать себя одиноким, если эти контакты поверхностны и не дают ощущения подлинной связи. Исследования показывают, что именно субъективное одиночество, а не объективная изоляция, является более сильным предиктором неблагоприятных последствий для здоровья. Это подчёркивает значимость не просто расширения социальной сети, но улучшения качества отношений — их глубины, взаимности, надёжности.
Негативные социальные взаимодействия — конфликты, критика, отвержение, предательство — оказывают непропорционально сильное воздействие на благополучие по сравнению с позитивными взаимодействиями. Исследования в области психологии отношений устойчиво обнаруживают асимметрию влияния: негативные события в отношениях влияют на удовлетворённость и здоровье сильнее, чем позитивные события сопоставимой интенсивности. Это соответствует общему принципу «негативного смещения» в психологии: негативная информация привлекает больше внимания, лучше запоминается, сильнее влияет на оценки и решения. Эволюционно это имеет смысл: угрозы требуют немедленного реагирования, тогда как возможности могут подождать. Однако в контексте отношений это означает, что единичный конфликт или эпизод критики может перевесить множество позитивных взаимодействий. Джон Готтман, изучавший супружеские отношения, обнаружил, что для стабильного и удовлетворительного брака необходимо соотношение позитивных и негативных взаимодействий примерно пять к одному — так называемое «магическое соотношение». Хронически конфликтные отношения, где негативные взаимодействия преобладают, становятся источником устойчивого стресса с соответствующими физиологическими последствиями.
Социальная поддержка реализуется через несколько различных функций, каждая из которых вносит свой вклад в буферирование стресса. Эмоциональная поддержка включает выражение заботы, сочувствия, принятия, подтверждение ценности человека и его чувств; она особенно важна при неконтролируемых стрессорах, где изменить ситуацию невозможно, но можно получить утешение и ощущение, что ты не один. Инструментальная поддержка предполагает конкретную помощь — деньгами, временем, услугами, — которая может уменьшить объективную тяжесть стрессовой ситуации. Информационная поддержка включает советы, рекомендации, информацию, которые помогают лучше понять ситуацию и найти способы её разрешения. Оценочная поддержка предоставляет обратную связь, помогающую человеку оценить свои усилия и скорректировать стратегию. Принадлежность к сообществу, разделяющему ценности и идентичность, даёт ощущение связи и смысла, которое само по себе является ресурсом. Различные типы поддержки могут быть более или менее эффективны в зависимости от характера стрессора: при контролируемых стрессорах особенно ценна инструментальная и информационная поддержка, при неконтролируемых — эмоциональная.
Воспринимаемая доступность поддержки часто оказывается более значимой для благополучия, чем реально полученная поддержка, что указывает на когнитивные механизмы буферного эффекта. Знание того, что в случае необходимости помощь будет доступна, меняет когнитивную оценку ситуации: потенциально угрожающие события воспринимаются как менее угрожающие, когда человек уверен, что не останется с ними один на один. Это объясняет, почему люди с надёжными социальными связями демонстрируют сниженную стресс-реактивность даже в ситуациях, где поддержка фактически не оказывается. Интересно, что реально полученная поддержка иногда ассоциирована с худшими, а не лучшими исходами. Это может объясняться тем, что получение поддержки активирует чувство зависимости и некомпетентности, или тем, что поддержка ищется преимущественно в наиболее тяжёлых ситуациях, создавая ложную корреляцию. Также важен способ оказания поддержки: навязчивая, контролирующая или некомпетентная поддержка может усугублять стресс вместо того, чтобы его смягчать. Эффективная поддержка отвечает потребностям реципиента, уважает его автономию и предоставляется в форме, которая не подрывает его самооценку.
Практические следствия понимания социального измерения стресса затрагивают как индивидуальные стратегии, так и общественную политику. На индивидуальном уровне это означает признание инвестиций в отношения как инвестиций в здоровье: поддержание и развитие значимых социальных связей, умение принимать и оказывать поддержку, способность разрешать конфликты конструктивно — всё это составляющие устойчивости к стрессу. Терапевтические интервенции, направленные на улучшение социального функционирования — тренинги социальных навыков, групповая терапия, семейная терапия, — эффективны отчасти именно потому, что усиливают социальную поддержку как буфер стресса. На общественном уровне признание одиночества как проблемы общественного здоровья ведёт к разработке программ по укреплению социальных связей — созданию общественных пространств для социального взаимодействия, поддержке волонтёрства и общественной активности, преодолению барьеров для социального участия пожилых людей и других уязвимых групп. Некоторые страны уже создали правительственные инициативы по борьбе с одиночеством, признавая его как фактор риска общественного здоровья.
5.2 Семья и близкие связи — первая линия поддержки и уязвимости
Семья занимает особое место среди социальных контекстов стресса, являясь одновременно первичной средой, в которой формируются базовые паттерны стресс-реактивности, и основным источником как поддержки, так и стресса на протяжении всей жизни. В отличие от других социальных связей, которые человек в значительной мере выбирает, семья происхождения дана ему изначально и формирует его задолго до того, как он способен критически оценивать этот опыт или искать альтернативы. Ранний семейный опыт буквально встраивается в биологию развивающегося организма, калибруя нейроэндокринные системы стресс-ответа, формируя нейронные цепи эмоциональной регуляции, закладывая имплицитные модели отношений. Эта биологическая «запись» раннего опыта объясняет его долгосрочное влияние: паттерны, сформированные в первые годы жизни, сохраняются на протяжении десятилетий и влияют на здоровье, отношения, психологическое функционирование во взрослом возрасте. Одновременно семья остаётся важнейшим контекстом социальной поддержки и стресса для взрослых: качество партнёрских и родительских отношений является одним из сильнейших предикторов благополучия, а семейные конфликты и дисфункции — одним из главных источников хронического стресса.
Концепция неблагоприятного детского опыта, известная под аббревиатурой ACE от английского «Adverse Childhood Experiences», систематизирует понимание влияния ранних стрессоров на долгосрочное здоровье. Исследование, проведённое Винсентом Фелитти и Робертом Андой в девяностых годах на более чем семнадцати тысячах участников, выявило сильную дозозависимую связь между количеством неблагоприятных детских переживаний — физическим, эмоциональным и сексуальным насилием, пренебрежением, дисфункцией домохозяйства — и риском широкого спектра заболеваний и проблем во взрослом возрасте. По сравнению с людьми без неблагоприятного детского опыта те, кто пережил четыре и более категории таких переживаний, имели в четыре-двенадцать раз повышенный риск алкоголизма, наркотической зависимости, депрессии, попыток самоубийства, а также значительно повышенный риск ишемической болезни сердца, рака, хронических заболеваний лёгких, переломов, заболеваний печени. Эти данные произвели революцию в понимании истоков здоровья и болезни, показав, что «детские» переживания имеют «взрослые» последствия через десятилетия. Распространённость неблагоприятного детского опыта оказалась высокой даже в благополучных популяциях: около двух третей участников оригинального исследования сообщили хотя бы об одной категории, около двенадцати процентов — о четырёх и более.
Биологические механизмы, связывающие ранний стресс с долгосрочными последствиями для здоровья, включают программирование стресс-систем, эпигенетические изменения, нейроструктурные перестройки. Гипоталамо-гипофизарно-надпочечниковая ось калибруется в раннем развитии в соответствии с характеристиками среды: если среда безопасна и предсказуема, ось настраивается на умеренную базовую активность с гибкой реакцией на острые стрессоры и эффективным восстановлением. Если среда хронически угрожающа или непредсказуема, ось может стать гиперреактивной — давая избыточный ответ на относительно небольшие стрессоры — или, напротив, притуплённой — с недостаточной реактивностью после длительного истощения. Оба паттерна дисрегуляции ассоциированы с неблагоприятными последствиями для здоровья. Эпигенетические исследования демонстрируют, что ранний стресс вызывает изменения в метилировании генов, регулирующих стресс-ответ, включая ген глюкокортикоидного рецептора. Эти эпигенетические модификации могут сохраняться на протяжении жизни и даже передаваться следующим поколениям. На нейроструктурном уровне ранний стресс ассоциирован с уменьшением объёма гиппокампа и префронтальной коры, увеличением объёма миндалины, изменениями в белом веществе мозга, нарушениями функциональной связности между областями, участвующими в эмоциональной регуляции.
Качество ранней привязанности — эмоциональной связи между ребёнком и ухаживающим взрослым — является ключевым фактором, определяющим развитие способности к регуляции стресса. Теория привязанности, основанная Джоном Боулби и экспериментально разработанная Мэри Эйнсворт, описывает, как паттерны раннего взаимодействия с опекуном формируют внутренние рабочие модели — имплицитные ожидания относительно себя, других и отношений. Безопасная привязанность формируется, когда опекун доступен, отзывчив и предсказуем: ребёнок усваивает, что он достоин заботы, другие люди надёжны, а мир в целом безопасен. Небезопасная привязанность — тревожная, избегающая или дезорганизованная — формируется при непоследовательном, отвергающем или пугающем поведении опекуна. Исследования показывают, что безопасная привязанность ассоциирована с более эффективной регуляцией стресса в детстве и во взрослой жизни: безопасно привязанные дети демонстрируют умеренную кортизоловую реактивность на стресс и быстрое восстановление, тогда как небезопасно привязанные дети показывают паттерны гипер- или гипореактивности. Важно, что привязанность — не только следствие темперамента ребёнка, но преимущественно результат поведения опекуна, что открывает возможности для интервенций.
Семья не перестаёт быть значимым контекстом стресса и поддержки во взрослой жизни, хотя характер её влияния меняется. Партнёрские отношения занимают центральное место в социальной жизни большинства взрослых и оказывают мощное влияние на здоровье и благополучие. Качество брака является одним из наиболее устойчивых предикторов субъективного благополучия: люди в удовлетворительных партнёрских отношениях демонстрируют более высокий уровень счастья, более низкий уровень депрессии и тревоги, лучшие показатели физического здоровья. Биологические механизмы включают буферирование стресс-ответа поддерживающим партнёром, более здоровый образ жизни в контексте стабильных отношений, позитивное влияние на иммунную функцию. Однако неудовлетворительные партнёрские отношения — конфликтные, холодные, контролирующие — оказывают противоположный эффект: хронический стресс супружеского конфликта ассоциирован с повышенным риском депрессии, сердечно-сосудистых заболеваний, нарушений иммунной функции. Лабораторные исследования демонстрируют, что даже кратковременный конфликтный разговор между супругами вызывает повышение кортизола, провоспалительных цитокинов, нарушение заживления экспериментальных ран.
Родительство представляет собой ещё один контекст, в котором семья одновременно является источником глубокого смысла и значительного стресса. Появление ребёнка радикально перестраивает жизнь родителей, предъявляя новые требования и ограничивая ресурсы — время, сон, финансы, автономию. Исследования показывают, что переход к родительству часто сопровождается снижением удовлетворённости браком, хотя этот эффект модерируется качеством отношений до рождения ребёнка, разделением родительских обязанностей, социальной поддержкой. Воспитание детей с особыми потребностями, поведенческими проблемами или хроническими заболеваниями является документированным источником хронического стресса для родителей, ассоциированным с повышенным риском депрессии, тревоги, соматических проблем. Уход за стареющими родителями — задача, с которой сталкивается всё больше людей среднего возраста по мере увеличения продолжительности жизни — представляет собой ещё один семейный стрессор с документированными последствиями для здоровья ухаживающих. Так называемое «поколение сэндвича», одновременно заботящееся о детях и о престарелых родителях, испытывает особую нагрузку.
Межпоколенческая передача стресса и его последствий представляет собой одну из наиболее значимых и тревожных граней семейной динамики. Стресс родителей влияет на детей через множество путей. Пренатальный путь включает влияние материнского стресса во время беременности на развивающийся плод: кортизол матери проникает через плаценту, и хронически повышенный уровень может влиять на развитие мозга и стресс-систем плода. Эпигенетический путь предполагает передачу стресс-индуцированных эпигенетических модификаций через половые клетки; хотя этот механизм лучше документирован на животных моделях, имеются данные и для человека. Поведенческий путь включает влияние стресса родителей на их родительское поведение: родители в состоянии хронического стресса менее чувствительны к сигналам ребёнка, более раздражительны, менее последовательны, что нарушает развитие привязанности и саморегуляции у ребёнка. Моделирование означает, что дети усваивают стратегии совладания, наблюдая за родителями: если родители справляются со стрессом через алкоголь, агрессию или избегание, дети могут перенять эти паттерны. Эмоциональный климат семьи, в которой родители находятся под хроническим стрессом, характеризуется напряжённостью, конфликтностью, непредсказуемостью, что само по себе является стрессором для детей.
Понимание семьи как контекста стресса имеет важные терапевтические и профилактические импликации. Семейная терапия и терапия пар направлены на улучшение качества отношений, разрешение конфликтов, развитие коммуникативных навыков, что может уменьшать хронический стресс, генерируемый дисфункциональными отношениями. Программы поддержки родительства, особенно для семей группы риска, направлены на улучшение качества ранних отношений между родителем и ребёнком, что может прерывать межпоколенческую передачу стресса и его последствий. Вмешательства на ранних этапах развития, такие как программа «Nurse-Family Partnership», предполагающая регулярные визиты медсестёр к матерям из группы риска с первой беременности до двухлетия ребёнка, демонстрируют долгосрочные положительные эффекты для здоровья и развития детей. На уровне политики признание значимости ранних лет для долгосрочного здоровья обосновывает инвестиции в программы раннего детства, поддержку семей с маленькими детьми, оплачиваемые родительские отпуска, качественный и доступный уход за детьми. Экономические анализы показывают, что такие инвестиции многократно окупаются за счёт улучшения здоровья и продуктивности населения в долгосрочной перспективе.
Концепция устойчивости семьи дополняет фокус на семейных стрессорах и дисфункциях вниманием к факторам, которые позволяют семьям успешно справляться с трудностями. Исследования семей, переживших серьёзные кризисы — потерю члена семьи, тяжёлую болезнь, экономические потрясения, войну и вынужденное переселение, — выявляют факторы, отличающие семьи, которые сохраняют функционирование и сплочённость, от тех, которые распадаются. К таким факторам относятся гибкость семейной системы, способность к адаптации и реорганизации при изменении обстоятельств; связность и поддержка между членами семьи; открытая и конструктивная коммуникация; общие убеждения и смыслы, позволяющие осмыслять трудности; навыки совместного решения проблем; связи с более широким социальным окружением и сообществом. Эти факторы устойчивости могут быть целенаправленно развиты через семейные интервенции, что открывает перспективы профилактической работы с семьями, сталкивающимися с потенциально травматическими обстоятельствами.
5.3 Рабочая среда и организационный стресс
Работа занимает центральное место в жизни большинства взрослых людей: среднестатистический работающий человек проводит на рабочем месте или в связи с работой порядка сорока и более часов в неделю на протяжении четырёх-пяти десятилетий трудовой жизни. Этот колоссальный объём времени и вовлечённости делает рабочую среду одним из ключевых контекстов, определяющих здоровье и благополучие. Работа может быть источником смысла, самореализации, социальной идентичности, структуры, финансовой безопасности — факторов, которые сами по себе защитны для здоровья. Однако работа также является одним из главных источников хронического стресса в современном обществе, и организационный стресс — стресс, порождаемый условиями и характером работы — ассоциирован с серьёзными последствиями для физического и психического здоровья работников. Выгорание, депрессия, тревожные расстройства, сердечно-сосудистые заболевания, нарушения сна, скелетно-мышечные проблемы — всё это документированные последствия хронического рабочего стресса. Понимание организационных факторов, генерирующих стресс, необходимо как для индивидуальных стратегий совладания, так и для организационных и политических интервенций, направленных на создание более здоровой рабочей среды.
Модель требований-контроля, или модель напряжённости труда, разработанная социологом Робертом Карасеком в семидесятых годах двадцатого века, является одной из наиболее влиятельных теоретических рамок для понимания рабочего стресса. Согласно этой модели, стресс на работе определяется сочетанием двух измерений: психологических требований работы — объёма, интенсивности, сложности, временно́го давления — и степени контроля работника над своей деятельностью — возможности принимать решения, использовать свои навыки, влиять на условия работы. Наиболее стрессогенной является комбинация высоких требований и низкого контроля — так называемая «напряжённая работа»: работник вынужден много и интенсивно работать, но не имеет возможности влиять на то, как, когда и в какой последовательности выполнять задачи. Классическими примерами являются работа на конвейере, в колл-центре, определённые позиции в сфере услуг. Напротив, работа с высокими требованиями, но и высоким контролем — «активная работа» — менее стрессогенна и может даже способствовать развитию и обучению. Работа с низкими требованиями и низким контролем — «пассивная работа» — ассоциирована со скукой, демотивацией и постепенной утратой навыков. Многочисленные эпидемиологические исследования подтвердили связь «напряжённой работы» с повышенным риском сердечно-сосудистых заболеваний, депрессии, тревожных расстройств.
Модель дисбаланса усилий и вознаграждения, разработанная социологом Йоханнесом Зигристом, дополняет модель Карасека вниманием к реципрокности в трудовых отношениях. Согласно этой модели, стресс возникает, когда усилия, вкладываемые работником, — время, энергия, навыки, ответственность — не получают соразмерного вознаграждения в форме денег, статуса, признания, карьерных возможностей, стабильности занятости. Этот дисбаланс нарушает фундаментальную норму социальной взаимности и воспринимается как несправедливость, порождая хронический стресс. Ситуация усугубляется, если работник характеризуется так называемой сверхприверженностью — чрезмерной вовлечённостью в работу, неспособностью отключиться, потребностью в одобрении, — что делает его более уязвимым к последствиям дисбаланса. Эмпирические исследования подтверждают, что дисбаланс усилий и вознаграждения является независимым предиктором сердечно-сосудистых заболеваний, депрессии, нарушений сна, даже после контроля традиционных факторов риска. Модель особенно релевантна в условиях современной экономики с её нестабильной занятостью, размыванием границ между работой и личной жизнью, интенсификацией труда.
Психосоциальные факторы рабочей среды, выходящие за рамки требований и контроля, также вносят вклад в организационный стресс. Социальная поддержка на рабочем месте — со стороны коллег и руководителей — является важным модератором связи между рабочими стрессорами и их последствиями. Карасек и Терелл дополнили оригинальную модель требований-контроля третьим измерением — социальной поддержкой, — создав модель требований-контроля-поддержки. Работа с высокими требованиями, низким контролем и низкой поддержкой — «изолированно напряжённая» — оказывается наиболее патогенной. Организационная справедливость — воспринимаемая честность процедур, распределения ресурсов, межличностного обращения — является независимым предиктором здоровья работников: несправедливое обращение порождает хронический стресс независимо от других характеристик работы. Ролевая неопределённость и ролевой конфликт — ситуации, когда работник не имеет ясного понимания своих обязанностей или получает противоречивые требования — также ассоциированы со стрессом. Нестабильность занятости и угроза увольнения оказывают самостоятельное влияние на здоровье, независимо от того, реализуется ли угроза.
Эмоциональный труд — необходимость регулировать свои эмоции и выражать определённые эмоции как часть рабочих обязанностей — представляет собой специфический источник стресса в профессиях сферы услуг. Концепция, введённая социологом Арли Хохшильд, описывает требование улыбаться и быть приветливым с клиентами независимо от своего реального эмоционального состояния, подавлять раздражение и негативные чувства, демонстрировать энтузиазм и заботу. Бортпроводники, продавцы, официанты, медицинские сёстры, учителя, работники колл-центров — все эти профессии требуют значительного эмоционального труда. Проблема возникает, когда между требуемыми и реально переживаемыми эмоциями существует устойчивое расхождение — эмоциональный диссонанс. Поверхностное разыгрывание эмоций, которых не чувствуешь, является более истощающим, чем глубокое, при котором работник действительно пытается вызвать в себе требуемое состояние. Хронический эмоциональный труд ассоциирован с эмоциональным истощением, деперсонализацией, выгоранием. Признание эмоционального труда как реальной работы, требующей признания и вознаграждения, является важным шагом к более здоровой организации труда в сфере услуг.
Выгорание представляет собой синдром, специфически связанный с хроническим рабочим стрессом, который не был успешно преодолён. Согласно определению Всемирной организации здравоохранения в Международной классификации болезней одиннадцатого пересмотра, выгорание характеризуется тремя измерениями: ощущением истощения или исчерпания энергии; нарастающей психической дистанцированностью от работы или чувствами негативизма и цинизма по отношению к работе; снижением профессиональной эффективности. Важно, что выгорание определено как профессиональный феномен, а не как медицинский диагноз, и отнесено к категории факторов, влияющих на состояние здоровья, а не к расстройствам. Выгорание особенно распространено в помогающих профессиях — медицине, образовании, социальной работе, — где эмоциональные требования высоки, а ресурсы часто ограничены. Пандемия показала масштаб проблемы: значительная доля медицинских работников во всём мире сообщала о симптомах выгорания. Последствия выгорания включают не только страдания самих работников, но и снижение качества предоставляемых услуг, ошибки, текучесть кадров, экономические потери для организаций и общества.
Связь организационного стресса с соматическим здоровьем документирована обширным корпусом эпидемиологических исследований, демонстрирующих, что рабочие условия являются детерминантами здоровья, а не только следствием индивидуальных характеристик работников. Исследование Уайтхолл, начатое Майклом Мармотом в семидесятых годах на британских государственных служащих, продемонстрировало социальный градиент здоровья даже среди работников с относительно благоприятными условиями труда: на каждой ступеньке служебной иерархии здоровье было хуже, чем на вышестоящей, и этот градиент объяснялся во многом различиями в контроле над работой. Метаанализы связывают напряжённую работу с повышением риска ишемической болезни сердца примерно на двадцать-пятьдесят процентов, с повышением риска инсульта, с увеличением риска диабета второго типа. Депрессия и тревожные расстройства также существенно чаще развиваются у работников в условиях хронического рабочего стресса. Нарушения сна, скелетно-мышечные боли, иммунная дисфункция дополняют картину влияния организационного стресса на здоровье. Экономические издержки включают затраты на лечение, потери продуктивности, абсентеизм и презентеизм, текучесть кадров, досрочный выход на пенсию.
Организационные и политические интервенции, направленные на снижение рабочего стресса, приобретают всё большее признание как необходимое дополнение к индивидуальным стратегиям совладания. Стандарт ИСО 45003, опубликованный в 2021 году, представляет собой первый международный стандарт, специально посвящённый управлению психосоциальными рисками на рабочем месте. Он предоставляет руководство по идентификации психосоциальных опасностей, оценке связанных рисков и разработке мер контроля, интегрированных в общую систему управления охраной труда. Организационные интервенции могут быть первичными — направленными на устранение или снижение источников стресса (редизайн работы, увеличение контроля и автономии, справедливое распределение нагрузки), вторичными — направленными на повышение способности работников справляться со стрессом (тренинги управления стрессом, программы поддержки работников), или третичными — направленными на помощь уже пострадавшим (лечение, реабилитация). Исследования показывают, что первичные, организационные интервенции более эффективны для улучшения здоровья работников, чем исключительно индивидуальные, хотя оптимальным является комбинированный подход.
Признание организационного стресса как проблемы общественного здоровья, а не только индивидуальной неспособности «справляться», представляет собой важный концептуальный сдвиг с практическими последствиями. Фокусировка на «стрессоустойчивости» отдельных работников, при игнорировании стрессогенных характеристик самой работы, перекладывает ответственность с организаций на индивидов и не решает проблему на системном уровне. Если значительная доля работников определённой организации или профессии демонстрирует симптомы выгорания, проблема, очевидно, не в недостаточной устойчивости каждого из них, а в условиях работы. Это не умаляет значимости индивидуальных стратегий совладания, но указывает на их недостаточность без организационных изменений. Трудовое законодательство, коллективные договоры, культура организации, управленческие практики — всё это определяет, насколько стрессогенной или, напротив, поддерживающей здоровье является рабочая среда. Политика, основанная на научных данных о рабочем стрессе, может существенно улучшить здоровье работающего населения и принести экономические выгоды через повышение продуктивности и снижение затрат, связанных с нездоровьем.
5.4 Культурный контекст — как общество формирует стресс
Культура представляет собой всеобъемлющий контекст, в котором разворачивается человеческая жизнь, и её влияние на переживание стресса столь глубоко, что часто остаётся незамеченным — подобно воде для рыбы, культура настолько пронизывает наш опыт, что мы редко осознаём её формирующее воздействие. Культура определяет, какие события считаются стрессорами, а какие — нормальной частью жизни; какие эмоциональные реакции приемлемы, а какие должны подавляться; какие стратегии совладания поощряются, а какие порицаются; к кому уместно обращаться за помощью и какого рода помощь считается приемлемой. Эти культурно обусловленные рамки не являются произвольными конструкциями — они отражают адаптацию сообществ к их экологическим, историческим и социальным условиям и передаются через поколения посредством социализации, языка, ритуалов, институтов. Однако те же культурные рамки, которые обеспечивают осмысленность и предсказуемость социальной жизни, могут становиться источником стресса, когда индивидуальный опыт не вписывается в культурно одобренные шаблоны или когда культурные ожидания вступают в конфликт друг с другом или с изменившимися обстоятельствами. Понимание культурного измерения стресса необходимо для адекватной интерпретации данных исследований, проведённых в различных культурных контекстах, и для разработки культурно компетентных интервенций.
Концептуализация стресса и связанных с ним понятий существенно различается между культурами, что создаёт как теоретические, так и практические сложности. Само понятие «стресс» в его современном значении является относительно недавним продуктом западной, преимущественно англоязычной научной традиции и не имеет прямых эквивалентов во многих других языках и культурных традициях. В некоторых культурах дистресс концептуализируется преимущественно в соматических терминах — как телесные симптомы и дисбаланс физиологических процессов, — тогда как в других акцент делается на эмоциональных или духовных аспектах. Культурно-специфические синдромы — такие как «нервы» в латиноамериканских культурах, «хва-бён» в корейской культуре или «суто» в культурах Латинской Америки — представляют собой способы выражения и осмысления дистресса, которые не полностью соответствуют западным диагностическим категориям, но являются осмысленными и функциональными в своих культурных контекстах. Исследователи должны быть осторожны, экстраполируя концепции и измерительные инструменты, разработанные в одной культуре, на другие, поскольку за внешним сходством симптомов могут скрываться различные смыслы и механизмы.
Различие между индивидуалистическими и коллективистскими культурами, хотя и представляет собой упрощение реального культурного разнообразия, остаётся полезной эвристикой для понимания культурных различий в переживании и совладании со стрессом. В культурах, которые принято относить к индивидуалистическим — преимущественно западных, особенно англосаксонских, — акцент делается на автономии, самореализации, личных достижениях, уникальности индивида. Стрессоры в таких культурах часто связаны с угрозами автономии, неудачами в личных достижениях, ограничениями свободы выбора. Ценимые стратегии совладания включают активное решение проблем, прямую коммуникацию, обращение к профессиональной помощи. В культурах, которые принято относить к коллективистским — многих азиатских, африканских, латиноамериканских, — акцент делается на гармонии в отношениях, выполнении социальных обязательств, принадлежности к группе. Стрессоры здесь чаще связаны с угрозами групповой гармонии, неспособностью выполнить обязательства перед семьёй или сообществом, потерей лица. Ценимые стратегии совладания могут включать принятие, терпение, непрямую коммуникацию, опору на семью и сообщество. Эти различия не являются абсолютными — внутри каждой культуры существует значительное разнообразие, и глобализация размывает традиционные культурные границы, — но они указывают на важность культурного контекста для понимания стресса.
Культурные нормы выражения эмоций существенно влияют на то, как стресс проявляется и как на него реагируют окружающие. Культуры различаются по степени, в которой открытое выражение эмоций — особенно негативных — считается приемлемым и даже желательным. В некоторых культурных контекстах выражение дистресса через плач, жалобы, открытое проявление тревоги или грусти рассматривается как естественное и здоровое, приглашающее социальную поддержку. В других контекстах те же проявления могут восприниматься как слабость, неуместность, нагрузка на окружающих; ценятся эмоциональная сдержанность, стоицизм, сохранение «лица». Эти нормы формируются с раннего детства через процессы социализации: родители и другие взрослые по-разному реагируют на эмоциональные проявления детей, поощряя одни и санкционируя другие. Последствия этих различий для здоровья неоднозначны: с одной стороны, подавление эмоций ассоциировано с физиологическими издержками; с другой стороны, эмоциональная сдержанность может быть функциональной в контексте, где открытое выражение несёт социальные риски. Ключевым является соответствие между индивидуальным стилем и культурным контекстом.
Стигма психических проблем и обращения за помощью варьирует между культурами и существенно влияет на то, получают ли люди, страдающие от последствий стресса, необходимую помощь. Во многих культурах психические проблемы стигматизированы сильнее, чем соматические заболевания: признание в тревоге или депрессии может восприниматься как постыдное признание слабости, неполноценности, потенциально порочащее не только самого человека, но и его семью. Обращение к психиатру или психотерапевту может восприниматься как признание «сумасшествия» и нести серьёзные социальные последствия — для репутации, карьеры, брачных перспектив. В таких контекстах люди чаще обращаются за помощью к врачам общей практики с соматическими жалобами, к традиционным целителям, к религиозным лидерам или справляются самостоятельно и с опорой на семью. Это не означает, что западная модель профессиональной психологической помощи является единственно правильной — традиционные системы поддержки могут быть весьма эффективны, — но указывает на важность культурной адаптации служб психического здоровья и работы по снижению стигмы.
Религия и духовность представляют собой важное измерение культуры, которое оказывает существенное влияние на переживание и совладание со стрессом. Религиозные верования предоставляют рамки для осмысления страдания, придавая ему значение и место в более широкой картине мира. Идея кармы, воли Божьей, испытания или очищения позволяет интегрировать болезненный опыт в осмысленный нарратив. Религиозные практики — молитва, медитация, ритуалы — могут обеспечивать регуляцию эмоций и физиологического состояния. Религиозные сообщества предоставляют социальную поддержку, принадлежность, идентичность. Эмпирические исследования связи религиозности и здоровья демонстрируют преимущественно положительные ассоциации, хотя механизмы сложны и включают как специфически религиозные факторы, так и опосредующие переменные — социальную поддержку общины, более здоровый образ жизни, чувство смысла и надежды. Однако религия может быть и источником стресса — когда порождает чувство вины, конфликт с научным мировоззрением, отвержение со стороны религиозного сообщества или когда используется для оправдания насилия и дискриминации. Понимание роли религии в жизни конкретного человека важно для культурно компетентной помощи.
Процессы аккультурации — адаптации к новой культуре при миграции или в ситуации культурного контакта — представляют собой специфический источник стресса, который приобретает всё большее значение в эпоху глобальной мобильности. Аккультурационный стресс возникает из необходимости адаптироваться к новому языку, социальным нормам, институтам, ожиданиям, часто при одновременной утрате привычной социальной поддержки, статуса, идентичности. Мигранты и беженцы сталкиваются с множеством стрессоров: языковые барьеры, дискриминация, неопределённость правового статуса, разлука с семьёй, утрата профессионального статуса, культурный шок. Исследования показывают, что аккультурационный стресс ассоциирован с повышенным риском депрессии, тревожных расстройств, соматических симптомов. Стратегии аккультурации различаются: интеграция — сохранение связи с культурой происхождения при одновременном освоении новой культуры — обычно ассоциирована с лучшими исходами, чем ассимиляция, сепарация или маргинализация. Однако возможность интеграции зависит не только от индивидуального выбора, но и от политики и отношения принимающего общества. Дети мигрантов, растущие между двумя культурами, сталкиваются со специфическими вызовами формирования идентичности.
Избегание культурных стереотипов и признание внутрикультурного разнообразия являются важными методологическими и этическими принципами при обсуждении культурных различий. Утверждения о том, что «азиаты справляются со стрессом так-то» или «латиноамериканцы переживают эмоции иначе», являются чрезмерными обобщениями, игнорирующими огромное разнообразие внутри каждой культурной группы. Индивидуальные различия внутри культуры часто превышают средние различия между культурами. Более того, культуры не являются статичными, гомогенными сущностями — они динамичны, внутренне разнородны, пересекаются друг с другом. Социальный класс, образование, городское или сельское происхождение, поколение, гендер, индивидуальная история — все эти факторы создают вариативность внутри любой культурной группы. Культурная компетентность означает не знание фиксированных «фактов» о различных культурах, а чувствительность к возможному культурному влиянию, готовность исследовать культурный контекст конкретного человека, рефлексию собственных культурных предубеждений. Это особенно важно для специалистов, работающих с культурно разнообразными популяциями.
Практические следствия понимания культурного контекста стресса затрагивают исследования, клиническую практику и политику. Для исследований это означает необходимость кросс-культурной валидации инструментов измерения стресса и связанных конструктов, осторожность в генерализации результатов, полученных в одной культуре, на другие, включение культурных переменных в исследовательские модели. Для клинической практики это означает культурную адаптацию интервенций — не просто перевод, но адаптацию содержания, формата, способа подачи к культурному контексту клиентов; учёт культурных объяснительных моделей болезни; работу с культурной идентичностью и аккультурационными проблемами; осознание собственной культурной позиционированности терапевта. Для политики это означает внимание к культурным потребностям различных групп населения при разработке программ и услуг, преодоление барьеров доступа для культурных меньшинств, поддержку культурного разнообразия как ресурса, а не проблемы. Культурная перспектива обогащает понимание стресса, напоминая, что универсальные биологические механизмы реализуются в культурно специфических формах.
5.5 Социально-экономический статус — неравенство как хронический стрессор
Социально-экономический статус — композитный показатель, включающий уровень образования, доход, профессию и связанные с ними ресурсы и престиж — является одним из наиболее мощных и устойчивых предикторов здоровья, обнаруживаемых в эпидемиологических исследованиях. Связь между социальным положением и здоровьем документирована для широчайшего спектра заболеваний и причин смерти, для разных стран, исторических периодов, методологий исследования. Люди с более низким социально-экономическим статусом живут в среднем меньше, болеют чаще и тяжелее, имеют худшее качество жизни, связанное со здоровьем. Это не новое открытие — связь бедности и болезней отмечалась ещё в девятнадцатом веке, — однако современные исследования существенно углубили понимание механизмов этой связи. Стресс занимает центральное место среди этих механизмов: низкий социально-экономический статус ассоциирован с большей подверженностью разнообразным стрессорам и с меньшими ресурсами для совладания с ними, что создаёт условия для хронического стресса с его биологическими последствиями. Понимание того, как социальное неравенство транслируется в неравенство здоровья через стрессовые механизмы, имеет критическое значение для общественного здравоохранения и социальной политики.
Важнейшим открытием современной социальной эпидемиологии является обнаружение социального градиента здоровья — того факта, что связь между статусом и здоровьем не ограничивается контрастом между бедностью и достатком, но представляет собой непрерывный градиент, охватывающий всю социальную иерархию. Исследования Уайтхолл, начатые Майклом Мармотом в семидесятых годах на когорте британских государственных служащих — популяции, в которой никто не живёт в бедности и все имеют стабильную занятость и доступ к медицине, — продемонстрировали, что на каждой ступеньке служебной иерархии смертность выше, чем на вышестоящей. Клерки умирали чаще, чем менеджеры среднего звена, те — чаще, чем высшие администраторы. Этот градиент сохранялся после контроля традиционных факторов риска — курения, холестерина, артериального давления, — что указывало на существование дополнительных механизмов. Аналогичные градиенты обнаружены во множестве других исследований и популяций. Это означает, что проблема не сводится к крайней бедности и её очевидным последствиям — недоеданию, отсутствию медицинской помощи, опасным условиям жизни; даже относительное положение в социальной иерархии влияет на здоровье, и этот эффект распространяется на всё население.
Низкий социально-экономический статус ассоциирован с повышенной экспозицией к разнообразным стрессорам, что создаёт условия для хронического стрессового воздействия. Финансовая нестабильность — невозможность оплатить неожиданные расходы, угроза потери жилья, долговое бремя — является источником постоянного беспокойства и требует непрерывных усилий по управлению ограниченными ресурсами. Жилищные условия могут включать переполненность, шум, загрязнение воздуха, отсутствие зелёных пространств, криминальное окружение — всё это документированные стрессоры с биологическими последствиями. Рабочие условия чаще характеризуются высокими требованиями и низким контролем, нестабильной занятостью, физическими опасностями, низким вознаграждением — теми самыми факторами организационного стресса, которые обсуждались ранее. Доступ к качественному питанию, медицинской помощи, образованию для детей, безопасному отдыху ограничен. Социальные отношения могут страдать от напряжений, связанных с финансовыми трудностями. Таким образом, низкий статус означает не один изолированный стрессор, но кумуляцию множества стрессоров, которые взаимодействуют и усиливают друг друга.
Концепция аллостатической нагрузки бедности описывает биологическое накопление последствий хронического стресса, связанного с социально-экономической депривацией. Аллостатическая нагрузка — совокупный износ физиологических систем вследствие хронической адаптации к стрессорам — измеряется через комплекс биомаркёров, включающих показатели сердечно-сосудистой системы, метаболизма, иммунной функции, нейроэндокринной регуляции. Исследования устойчиво демонстрируют, что люди с низким социально-экономическим статусом имеют более высокие показатели аллостатической нагрузки даже в относительно молодом возрасте, что указывает на ускоренное биологическое старение. Этот эффект наблюдается уже у детей и подростков из неблагополучных семей, что свидетельствует о раннем начале биологической «записи» социального неравенства. Аллостатическая нагрузка предсказывает последующее развитие хронических заболеваний и преждевременную смертность, что делает её ценным интегративным показателем кумулятивного влияния социального неблагополучия на здоровье. Таким образом, социальное неравенство буквально воплощается в теле, преобразуясь из социального факта в биологическую реальность.
Субъективное восприятие социального положения — того, где человек видит себя на социальной лестнице относительно других — оказывается важным предиктором здоровья, иногда даже более сильным, чем объективные показатели статуса. Это указывает на психологические механизмы связи статуса и здоровья, выходящие за пределы материальных условий. Осознание собственного низкого положения, ощущение подчинённости, неуважения, несправедливости порождает хронический стресс независимо от абсолютного уровня ресурсов. Социальное сравнение — сравнение себя с более успешными другими — может быть источником дистресса, особенно в обществах с высоким неравенством и культурой, акцентирующей индивидуальный успех. Исследования на животных моделях демонстрируют, что низкий ранг в социальной иерархии ассоциирован со стрессовыми биологическими профилями — повышенным кортизолом, воспалением, атеросклерозом — даже при контроле материальных условий, что подтверждает самостоятельное значение социальной позиции. У людей субъективный статус ассоциирован с теми же биомаркёрами стресса и с показателями здоровья. Это объясняет, почему неравенство как характеристика общества, а не только индивидуальная бедность, связано с худшим здоровьем населения.
Дискриминация представляет собой особый механизм, через который социальное неблагополучие транслируется в хронический стресс, особенно для членов маргинализированных групп — расовых и этнических меньшинств, женщин, людей с инвалидностью, сексуальных меньшинств. Переживание дискриминации — явной или скрытой, институциональной или межличностной — является документированным стрессором с биологическими последствиями. Исследования афроамериканцев в США демонстрируют, что переживаемая расовая дискриминация ассоциирована с повышенным артериальным давлением, хроническим воспалением, ускоренным клеточным старением, повышенным риском сердечно-сосудистых заболеваний, негативными исходами беременности. Концепция «атмосферного воздействия» описывает кумулятивный эффект повторяющихся стрессоров дискриминации на протяжении жизни, который «изнашивает» организм подобно погоде, разрушающей фасад здания. Стресс меньшинств — специфический хронический стресс, переживаемый членами стигматизированных групп вследствие их статуса меньшинства — включает не только дискриминационные события, но и ожидание дискриминации, необходимость скрывать идентичность, усвоенные негативные установки. Этот механизм частично объясняет диспропорции в здоровье между группами.
Детская бедность заслуживает особого внимания, поскольку её последствия для здоровья особенно глубоки и долгосрочны вследствие биологической чувствительности раннего развития к средовым воздействиям. Дети, растущие в бедности, подвергаются множественным стрессорам: нестабильность жилья и питания, конфликты в семье, часто связанные с финансовым стрессом родителей, проживание в неблагополучных районах, более ограниченный доступ к качественному образованию и медицине. Хронический стресс в критические периоды развития мозга влияет на формирование нейронных цепей, участвующих в эмоциональной регуляции и когнитивных функциях. Эпигенетические изменения, индуцированные ранним стрессом, могут сохраняться на протяжении жизни. Как результат, детская бедность предсказывает худшее здоровье и более низкий социально-экономический статус во взрослом возрасте, создавая межпоколенческий цикл неравенства. Эти данные подчёркивают критическую важность инвестиций в ранние годы жизни и в поддержку семей с детьми как стратегии улучшения здоровья населения и разрыва циклов депривации.
Структурный характер социально-экономического стресса требует структурных решений, выходящих за рамки индивидуальных вмешательств. Хотя индивидуальные стратегии совладания со стрессом, программы укрепления психического здоровья, медицинские интервенции важны и полезны, они не могут устранить корневые причины социально-экономических диспропорций в здоровье. Если источник стресса — финансовая нестабильность, плохие жилищные условия, нестабильная занятость, дискриминация, — то устранение этих источников требует изменений в экономической политике, трудовом законодательстве, жилищной политике, антидискриминационных мерах. Перенос ответственности за здоровье исключительно на индивидов — призывы «управлять стрессом», «вести здоровый образ жизни» — при игнорировании структурных барьеров может даже усугублять проблему, добавляя чувство вины и несостоятельности к уже имеющемуся стрессу. Эффективная политика здоровья должна признавать социальные детерминанты здоровья и воздействовать на них. Примеры эффективных интервенций включают программы гарантированного минимального дохода, доступное жильё, качественные ранние образовательные программы, политику занятости, обеспечивающую достойные условия труда.
Понимание социально-экономического статуса как детерминанты здоровья через механизм стресса имеет глубокие импликации для науки о стрессе в целом. Оно напоминает, что стресс — не только индивидуальный, психологический феномен, но и социально-структурное явление, укоренённое в распределении ресурсов, власти, возможностей в обществе. Изучение стресса, ограниченное лабораторными экспериментами или клиническими популяциями, упускает этот структурный уровень. Интервенции, направленные на улучшение индивидуального совладания, не заменяют политики, направленной на снижение неравенства. Справедливость — не только этический императив, но и фактор общественного здоровья: более равные общества имеют лучшие показатели здоровья населения, включая психическое здоровье. Наука о стрессе, осведомлённая о социальных детерминантах, может вносить вклад в обоснование политики, направленной на создание более здорового и справедливого общества.
5.6 Общественные системы и политика здоровья
Уровень анализа, выходящий за пределы межличностных отношений, организаций и даже социально-экономической структуры, охватывает общественные институты и политические системы, которые формируют условия жизни целых популяций и, соответственно, их подверженность стрессу и его последствиям. Системы образования, здравоохранения, социальной защиты, трудовое законодательство, городское планирование, экологическая политика — все эти институциональные структуры создают контекст, в котором разворачивается повседневная жизнь людей, и определяют, какие стрессоры будут присутствовать в этой жизни и какие ресурсы для совладания будут доступны. Признание этого уровня влияния представляет собой радикальное расширение перспективы науки о стрессе: от фокуса на индивидуальных различиях в реактивности и совладании к пониманию структурных условий, которые генерируют или предотвращают стресс на уровне населения. Это расширение имеет критически важные практические следствия: если стресс существенно определяется политикой, то и его предотвращение требует политических решений, а наука о стрессе должна информировать эти решения.
Государственная политика в области социальной защиты оказывает измеримое влияние на здоровье и благополучие населения, и страны существенно различаются по степени этой защиты. Оплачиваемые отпуска по уходу за ребёнком позволяют родителям посвятить критические первые месяцы формированию привязанности без финансового стресса; страны с более щедрыми родительскими отпусками демонстрируют лучшие показатели материнского и детского здоровья. Пособия по безработице смягчают стресс потери работы, предоставляя время для поиска нового места без немедленного финансового краха; более щедрые пособия ассоциированы с лучшими показателями психического здоровья безработных. Пенсионное обеспечение влияет на благополучие пожилых людей и их семей. Доступность жилья определяет, будет ли жилищная неопределённость источником хронического стресса. Минимальная заработная плата и трудовое законодательство влияют на условия труда и финансовую безопасность работающих. Сравнительные исследования различных стран и естественные эксперименты — ситуации, когда политика изменяется, позволяя оценить последствия, — демонстрируют причинное влияние этих политических мер на показатели здоровья.
Система здравоохранения и доступность медицинской помощи имеют очевидное значение для здоровья населения, однако их влияние на стресс менее очевидно и заслуживает рассмотрения. Неуверенность в возможности получить медицинскую помощь при необходимости — из-за отсутствия страховки, недоступности услуг, высокой стоимости — сама по себе является источником хронического стресса, особенно для людей с хроническими заболеваниями или семей с детьми. Финансовый стресс, связанный с медицинскими расходами, является документированной проблемой в странах без универсального медицинского покрытия. Качество первичной медицинской помощи влияет на раннее выявление и лечение стресс-связанных расстройств. Доступность служб психического здоровья определяет, получат ли люди, страдающие от последствий стресса, своевременную и качественную помощь. Интеграция физической и психической помощи — понимание того, что психическое и физическое здоровье неразделимы — важна для холистического подхода к стресс-связанным состояниям. Профилактическая ориентация системы здравоохранения, в отличие от исключительно курационной, может предотвращать развитие стресс-связанных заболеваний.
Городская среда и жилищная политика формируют повседневный контекст жизни большинства населения в урбанизированных обществах и существенно влияют на стресс и здоровье. Качество жилья — теплоизоляция, вентиляция, отсутствие плесени и вредителей, адекватное пространство — влияет на физическое здоровье и психологическое благополучие. Характеристики района — уровень преступности, социальная сплочённость, доступность услуг — определяют социальный контекст жизни. Экологические факторы городской среды — качество воздуха, шум, доступность зелёных пространств — оказывают документированное влияние на стресс и здоровье. Исследования демонстрируют, что проживание вблизи зелёных пространств ассоциировано со сниженным уровнем кортизола, лучшим психическим здоровьем, большей физической активностью. Шум транспорта и другие источники шумового загрязнения являются стрессорами с биологическими последствиями — нарушениями сна, повышением артериального давления, сердечно-сосудистыми рисками. Городское планирование, учитывающее воздействие среды на здоровье, может создавать более здоровые города через зонирование, развитие общественного транспорта, создание парков, снижение загрязнения.
Система образования влияет на здоровье не только через результаты — более образованные люди, как правило, более здоровы, — но и через сам процесс образования и условия, в которых он происходит. Школьная среда может быть источником как поддержки, так и значительного стресса для детей и подростков: академическое давление, социальные отношения со сверстниками, буллинг, отношения с учителями — всё это влияет на психологическое благополучие и формирующиеся системы стресс-ответа. Школы, осознающие свою роль в укреплении психического здоровья, могут интегрировать программы социально-эмоционального обучения, создавать поддерживающий климат, обучать навыкам совладания со стрессом. Высшее образование сопряжено со специфическими стрессорами — академическим давлением, финансовой нестабильностью у студентов, неопределённостью будущего — и проблема психического здоровья студентов приобретает всё большее признание. Политика в области образования — размер классов, подготовка учителей, внимание к благополучию учащихся, а не только к академическим результатам — может влиять на стресс и здоровье целых поколений.
Рабочая политика, выходящая за рамки отдельных организаций на уровень национального законодательства и международных стандартов, определяет условия труда для миллионов работников. Регулирование рабочего времени — максимальная продолжительность рабочей недели, обязательные перерывы, ограничения сверхурочной работы — защищает от чрезмерной нагрузки. Законодательство о безопасности труда, включая признание психосоциальных рисков наряду с физическими, обязывает работодателей заботиться о психическом здоровье работников. Защита от увольнения и гарантии занятости снижают стресс неопределённости. Право на коллективные переговоры позволяет работникам влиять на условия труда. Баланс работы и личной жизни поддерживается политикой в области родительских отпусков, права на гибкий график, ограничения ожидаемой доступности вне рабочего времени. Международная организация труда и другие международные органы разрабатывают стандарты и руководства, однако их реализация зависит от национальной политики и правоприменения.
Экологическая политика и проблема изменения климата приобретают всё большее значение как факторы общественного здоровья, включая психическое здоровье и стресс. Загрязнение окружающей среды — воздуха, воды, почвы — оказывает как прямое токсическое воздействие на здоровье, так и косвенное через стресс обеспокоенности и неопределённости. Изменение климата генерирует стресс через множество путей: непосредственное воздействие экстремальных погодных явлений — наводнений, засух, волн жары; постепенные изменения среды обитания и средств к существованию; вынужденная миграция; неопределённость будущего; феномен «экологической тревоги» или «климатического горя» — психологический дистресс, связанный с осознанием экологического кризиса. Наиболее уязвимые группы — бедные сообщества, коренные народы, развивающиеся страны — несут непропорциональное бремя климатического стресса. Политика смягчения изменения климата и адаптации к нему имеет, таким образом, значение и для психического здоровья. Концепция «планетарного здоровья» интегрирует понимание того, что здоровье человека неотделимо от здоровья планеты.
Признание того, что стресс является не только индивидуальной, но и общественной проблемой, требующей политических решений, представляет собой важный концептуальный сдвиг с практическими последствиями. Традиционный подход к стрессу, фокусирующийся на индивидуальных различиях, личной ответственности, техниках самопомощи, является необходимым, но недостаточным. Если значительная доля стресса генерируется социальными условиями — бедностью, неравенством, прекарной занятостью, дискриминацией, неадекватными социальными услугами, — то индивидуальные вмешательства не могут решить проблему на популяционном уровне. Политика, основанная на научных данных о стрессе и его детерминантах, может улучшить здоровье миллионов людей через создание условий, предотвращающих хронический стресс или смягчающих его последствия. Это не умаляет значимости индивидуальной устойчивости и личных ресурсов, но дополняет её вниманием к структурным условиям, которые делают устойчивость более или менее необходимой и достижимой.
Оптимистическое заключение темы социального уровня анализа состоит в признании того, что социальные условия, генерирующие стресс, являются не природной данностью, а результатом человеческих решений и, следовательно, поддаются изменению. История общественного здравоохранения демонстрирует, что структурные изменения — улучшение санитарии, регулирование рабочих условий, развитие социальной защиты — приносили масштабные улучшения здоровья населения, часто более значительные, чем индивидуальные медицинские вмешательства. Современное понимание стресса как биопсихосоциального феномена, укоренённого в социальных условиях, предоставляет научную основу для политики, направленной на создание менее стрессогенного общества. Доказательная база для связи социальных условий со здоровьем через стрессовые механизмы достаточно убедительна для обоснования политических действий. Наука о стрессе, таким образом, выходит за пределы чисто академического предприятия и приобретает значение для социальной справедливости и общественного благополучия. Эта перспектива будет развита в последующих модулях курса, посвящённых контекстам и интервенциям на разных уровнях.

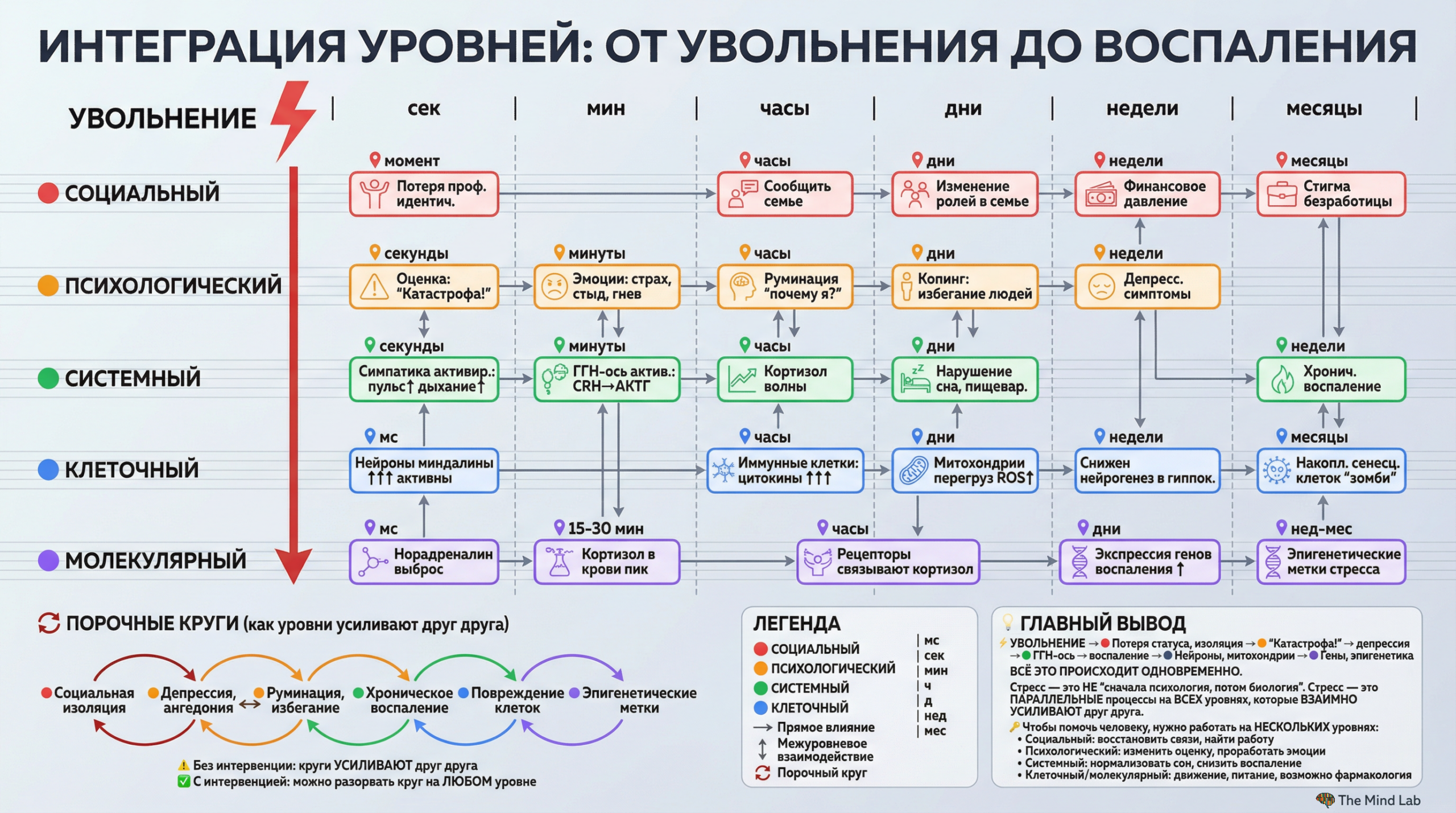
Академический слой
Перейти к практикуму1. Редукционизм и эмерджентность: философские основания многоуровневого анализа стресса
1.1 Редукционизм как научная стратегия и её успехи
Редукционистский подход представляет собой одну из наиболее влиятельных и продуктивных эпистемологических стратегий в истории естественных наук, и его значение для становления современной науки о стрессе невозможно переоценить. Сущность редукционизма состоит в методологическом принципе, согласно которому сложные явления могут и должны быть объяснены через свойства и взаимодействия их более простых составляющих. В контексте биологических и медицинских наук это означает стремление понять функционирование организма через изучение органов, органов через ткани, тканей через клетки, клеток через молекулы, а молекул через атомарные и субатомарные взаимодействия. Данная интеллектуальная традиция восходит к механистической философии семнадцатого века и получила мощное развитие в двадцатом столетии благодаря революционным открытиям в области молекулярной биологии, генетики и нейронаук. Применительно к изучению стресса редукционистская программа позволила трансформировать расплывчатое понятие о телесном напряжении в точную науку о конкретных молекулярных каскадах, рецепторных взаимодействиях и клеточных механизмах адаптации.
Исторический успех редукционизма в науке о стрессе наиболее ярко проявляется в расшифровке гипоталамо-гипофизарно-надпочечниковой оси, которая представляет собой центральный нейроэндокринный механизм стрессового ответа. До применения редукционистских методов представления о связи между психологическим переживанием и телесными изменениями оставались на уровне общих наблюдений и умозрительных теорий. Последовательное разложение этой сложной системы на составляющие элементы позволило идентифицировать кортиколиберин как гипоталамический пептид, инициирующий каскад, адренокортикотропный гормон как гипофизарный посредник, кортизол как конечный эффекторный гормон надпочечников. Каждый из этих компонентов был выделен, очищен, определена его химическая структура, синтезированы аналоги и антагонисты, установлены рецепторы и сигнальные пути. Эта кропотливая работа по декомпозиции системы на элементы создала основу для современного понимания того, как абстрактное переживание угрозы преобразуется в конкретные биохимические события, поддающиеся измерению и модификации.
Методологическая мощь редукционизма определяется несколькими фундаментальными преимуществами, которые делают его незаменимым инструментом научного познания. Первое и наиболее существенное преимущество связано с возможностью создания контролируемых экспериментальных условий, в которых исследователь может изолировать интересующую переменную от влияния побочных факторов. Когда клетка извлекается из организма и помещается в культуральную среду, исследователь получает возможность манипулировать концентрацией отдельных веществ, измерять ответы с высокой точностью и устанавливать причинно-следственные отношения между воздействием и эффектом. Второе преимущество состоит в достижении количественной точности и воспроизводимости результатов, что является краеугольным камнем научного метода. Третье преимущество связано с возможностью выявления универсальных механизмов, действующих на фундаментальном уровне организации живой материи. Открытие того, что глюкокортикоидные рецепторы функционируют по сходным принципам в клетках разных тканей и даже разных видов организмов, стало возможным именно благодаря редукционистскому анализу.
Расшифровка молекулярных механизмов рецепторной сигнализации представляет собой ещё один выдающийся пример успеха редукционистской программы в науке о стрессе. Понимание того, что гормоны стресса оказывают свои эффекты через связывание со специфическими белками-рецепторами, открыло совершенно новые горизонты для исследования и терапевтического вмешательства. Детальный анализ структуры глюкокортикоидного рецептора выявил его модульную организацию: лиганд-связывающий домен, ДНК-связывающий домен, трансактивационные регионы. Каждый из этих модулей был изучен отдельно с использованием методов рентгеновской кристаллографии, сайт-направленного мутагенеза, биохимического анализа. Это позволило понять, как связывание кортизола вызывает конформационные изменения рецептора, как рецептор перемещается в ядро клетки, как он взаимодействует с определёнными последовательностями ДНК и регулирует транскрипцию сотен генов-мишеней. Без такой детальной декомпозиции было бы невозможно разработать селективные модуляторы глюкокортикоидных рецепторов, которые сохраняют терапевтические эффекты при минимизации побочных.
Редукционистский подход оказался исключительно плодотворным и в области нейробиологии стресса, где он позволил картировать нейронные цепи, опосредующие различные компоненты стрессового ответа. Идентификация миндалевидного тела как ключевой структуры для детекции угрозы и формирования страха стала возможной благодаря систематическим исследованиям с использованием методов локальных повреждений, электрофизиологической регистрации и нейровизуализации. Дальнейшее разложение миндалины на ядра с различными функциями, а ядер на популяции нейронов с различными молекулярными маркерами и проекциями создало детальную карту того, как сенсорная информация об угрозе обрабатывается и преобразуется в поведенческие и вегетативные ответы. Современные методы оптогенетики и хемогенетики довели редукционистскую логику до предела, позволяя активировать или подавлять отдельные популяции нейронов с миллисекундной точностью и наблюдать последствия такой манипуляции для поведения и физиологии.
Фармакологический прогресс в лечении стресс-связанных расстройств является, пожалуй, наиболее практически значимым достижением редукционистской науки. Понимание роли серотониновой нейротрансмиссии в регуляции настроения и тревоги привело к разработке селективных ингибиторов обратного захвата серотонина, которые на протяжении десятилетий остаются препаратами первой линии при депрессии и тревожных расстройствах. Выявление роли гамма-аминомасляной кислоты как главного тормозного нейромедиатора центральной нервной системы легло в основу создания бензодиазепиновых анксиолитиков. Открытие глутаматергической дисфункции при стресс-связанных расстройствах открыло путь к принципиально новым терапевтическим подходам, включая использование кетамина при резистентной депрессии. Каждый из этих терапевтических прорывов стал возможным благодаря последовательному применению редукционистской логики: идентификация ключевой молекулы, понимание её синтеза, высвобождения, рецепторного связывания и инактивации, поиск соединений, модифицирующих эти процессы.
Эпистемологическое обоснование редукционизма опирается на фундаментальное представление о том, что законы природы иерархически организованы, и закономерности более высоких уровней в конечном счёте выводимы из закономерностей более низких уровней. Эта позиция, известная как теоретический или онтологический редукционизм, предполагает, что биологические явления в принципе объяснимы через химию, химия через физику, а все научные дисциплины образуют единую пирамиду с фундаментальной физикой в основании. Независимо от того, насколько полно эта программа реализуема на практике, она служит мощным эвристическим принципом, направляющим исследовательские усилия к поиску всё более глубоких механистических объяснений. В науке о стрессе этот принцип проявляется в стремлении объяснить психологические феномены через нейробиологию, нейробиологию через клеточную физиологию, клеточную физиологию через молекулярные взаимодействия. Даже если полная редукция недостижима, движение в этом направлении генерирует ценные знания о механизмах.
Важно подчеркнуть, что успехи редукционизма создали концептуальную и методологическую основу, на которой строится весь данный курс. Модули, посвящённые биологическим уровням анализа стресса, от молекулярного до системного, в значительной мере представляют собой изложение достижений редукционистской исследовательской программы. Понимание работы гипоталамо-гипофизарно-надпочечниковой оси в третьем модуле, механизмов нейропластичности во втором модуле, воспалительных каскадов в четвёртом модуле было бы невозможно без систематического применения редукционистских методов на протяжении десятилетий. Вместе с тем, как будет показано в последующих разделах, признание заслуг редукционизма не означает его абсолютизации. Понимание ограничений редукционистского подхода столь же важно для формирования адекватного научного мышления, как и понимание его достоинств, и именно этому посвящён следующий раздел.
1.2 Ограничения методологического редукционизма
При всей своей продуктивности редукционистская стратегия обнаруживает фундаментальные ограничения, когда применяется к изучению стресса как целостного биопсихосоциального феномена, и осознание этих ограничений составляет необходимое условие для формирования зрелого научного мышления. Первое и наиболее очевидное ограничение связано с неизбежной потерей контекста при изоляции объекта исследования от его естественного окружения. Когда нейрон извлекается из мозга и помещается в культуральную чашку, он лишается тысяч синаптических контактов с другими нейронами, утрачивает глиальное окружение, теряет связь с циркулирующими гормонами и иммунными медиаторами. То, что исследователь наблюдает в таких условиях, представляет собой поведение изолированной клетки, которое может существенно отличаться от её функционирования в составе интактной нервной системы. Парадокс редукционизма состоит в том, что стремление к экспериментальному контролю, достигаемое через изоляцию, одновременно разрушает тот самый контекст взаимодействий, который определяет реальное поведение системы.
Проблема потери контекста приобретает особую остроту в науке о стрессе, поскольку стрессовый ответ по своей природе является интегративным феноменом, возникающим из координированного взаимодействия множества систем организма. Кортизол, изучаемый в пробирке, демонстрирует определённые биохимические свойства, но его реальные эффекты в организме зависят от состояния нервной системы, уровня других гормонов, воспалительного статуса, циркадной фазы, пищевого статуса и множества других факторов. Рецептор, изучаемый в рекомбинантной системе экспрессии, показывает одни параметры связывания лиганда, но в контексте живой клетки его функция модулируется шаперонами, корегуляторами, пострансляционными модификациями и хроматиновым окружением генов-мишеней. Эти контекстуальные влияния не являются случайным шумом, который можно игнорировать; они составляют существенную часть того, как система реально функционирует. Редукционистский эксперимент, устраняя эти влияния ради чистоты, рискует изучать артефакт, а не реальность.
Второе фундаментальное ограничение редукционизма связано с проблемой временной динамики. Классический редукционистский эксперимент представляет собой срез в определённый момент времени: измеряется концентрация вещества, активность фермента, уровень экспрессии гена в данной точке. Однако стресс принципиально является процессом, разворачивающимся во времени, с различными фазами, переходами, петлями обратной связи и отсроченными эффектами. Острый стрессор вызывает волну изменений, которая распространяется через организм на протяжении минут, часов и дней, причём ранние изменения запускают поздние, а поздние модифицируют ответ на последующие стрессоры. Хронический стресс включает процессы адаптации, сенситизации, аллостатической перегрузки, которые разворачиваются на протяжении недель, месяцев и лет. Моментальный срез, каким бы детальным он ни был на молекулярном уровне, не схватывает эту динамическую сущность стрессового ответа. Понимание стресса требует отслеживания траекторий изменения во времени, что представляет существенно более сложную методологическую задачу.
Третье критическое ограничение редукционизма обусловлено фундаментальной нелинейностью биологических систем. Редукционистская логика имплицитно предполагает, что понимание частей позволит понять целое путём их суммирования, что эффект будет пропорционален причине, что малые изменения на входе будут вызывать малые изменения на выходе. Однако живые системы характеризуются нелинейными зависимостями, пороговыми эффектами, положительными обратными связями, которые нарушают эти простые отношения. В контексте стресса это проявляется множеством способов: длительный хронический стресс может не вызывать видимых нарушений до момента, когда система внезапно декомпенсируется; малое изменение в когнитивной оценке ситуации может радикально изменить весь паттерн биологического ответа; взаимодействие множества генетических вариантов, каждый из которых имеет ничтожный индивидуальный эффект, может создавать качественно различные фенотипы уязвимости к стрессу. Эти нелинейные эффекты возникают из взаимодействий между компонентами и не могут быть предсказаны из изучения компонентов по отдельности.
Многочисленные неудачи трансляции редукционистских открытий в клиническую практику служат красноречивой иллюстрацией обсуждаемых ограничений. Десятилетия исследований воспалительных механизмов депрессии выявили повышенные уровни провоспалительных цитокинов, таких как интерлейкин-6 и фактор некроза опухоли альфа, у значительной части пациентов с депрессией. Редукционистская логика предсказывала, что блокирование этих цитокинов должно облегчать депрессивную симптоматику. Клинические испытания антицитокиновых препаратов, однако, показали разочаровывающие результаты: эффекты были слабыми, нестабильными или отсутствовали. Аналогичная история повторялась с антагонистами рецепторов кортиколиберина, ингибиторами синтеза кортизола, модуляторами глутаматных рецепторов и многими другими соединениями, которые блокировали конкретные молекулярные мишени, идентифицированные редукционистскими исследованиями. Эти неудачи свидетельствуют не о том, что молекулярные механизмы нерелевантны, а о том, что изолированное воздействие на один компонент сложной системы не эквивалентно модификации функции системы в целом.
Концептуальная проблема декомпозируемости представляет ещё одно серьёзное ограничение редукционистской программы. Редукционизм предполагает, что систему можно разобрать на части, изучить каждую часть отдельно, а затем мысленно собрать обратно, получив понимание целого. Однако не все системы декомпозируемы в этом смысле. Если свойства системы существенно зависят от способа организации и взаимодействия её компонентов, то разборка системы уничтожает именно то, что мы пытаемся понять. Живой организм не является механизмом вроде часов, который можно разобрать и собрать; это динамическая система, непрерывно поддерживающая себя через потоки энергии и информации. Стрессовый ответ не локализован ни в одной молекуле, ни в одной клетке, ни в одном органе; он существует как паттерн координированных изменений, распределённый по всему организму. Попытка локализовать такой паттерн редукцией к элементам подобна попытке найти мелодию, разбирая музыкальный инструмент на части.
Ограничения редукционизма становятся особенно явными при попытке объяснить психологические феномены стресса через нейробиологические механизмы. Когнитивная оценка угрозы, субъективное переживание тревоги, волевое усилие по регуляции эмоций, смысловая переработка травматического опыта — все эти психологические процессы, несомненно, реализуются через активность нейронных сетей мозга. Однако редукция психологического описания к нейробиологическому сталкивается с проблемой уровней объяснения. Даже если бы мы знали состояние каждого нейрона в мозге человека, переживающего тревогу, это не дало бы нам того понимания, которое обеспечивает психологический анализ содержания его переживаний, истории формирования его страхов, стратегий совладания, которые он использует. Нейробиологическое и психологическое описания не являются взаимозаменяемыми; они отвечают на разные вопросы и служат разным объяснительным целям.
Осознание ограничений редукционизма не означает отказа от редукционистских методов, которые остаются незаменимыми инструментами научного познания. Речь идёт о понимании границ применимости этих методов и необходимости дополнения их другими подходами. В контексте данного курса это означает, что изучение молекулярных, клеточных и системных механизмов стресса в первых четырёх модулях должно сопровождаться пониманием того, что эти механизмы не исчерпывают феномен стресса. Психологические процессы, рассматриваемые в пятом модуле, социальные контексты, анализируемые в десятом модуле, не сводятся к биологическим основаниям, хотя и реализуются через них. Интегративное понимание стресса требует способности мыслить одновременно на нескольких уровнях, связывая их между собой, но не редуцируя один к другому. Такой способ мышления предполагает признание реальности эмерджентных свойств, которым посвящён следующий раздел.
1.3 Эмерджентность: качественные скачки между уровнями организации
Концепция эмерджентности представляет собой философский и научный ответ на ограничения редукционизма, предлагая понимание того, как сложные системы могут обладать свойствами, не присущими их составляющим элементам и не выводимыми из них простым суммированием. Сам термин происходит от латинского глагола, означающего «появляться» или «возникать», и указывает на феномен появления качественно новых характеристик при переходе от более простого уровня организации к более сложному. В контексте науки о стрессе эмерджентность проявляется на каждом переходе между уровнями анализа: от молекул к клеткам, от клеток к тканям и органам, от органов к системам организма, от биологических систем к психологическим процессам, от индивидуальной психологии к социальным феноменам. На каждом из этих переходов возникают свойства, которые невозможно обнаружить на предыдущем уровне, сколь бы детально он ни был изучен. Признание реальности эмерджентности имеет глубокие последствия для понимания природы стресса и стратегий его изучения.
Классические примеры эмерджентности в физике и химии помогают прояснить суть этого понятия прежде, чем обратиться к его проявлениям в биологии стресса. Отдельные молекулы воды не обладают свойствами жидкости: они не мокрые, не текучие, не имеют поверхностного натяжения. Эти свойства возникают только при объединении огромного числа молекул и представляют собой характеристики ансамбля, а не его элементов. Температура не является свойством отдельной молекулы; она возникает как статистическая мера средней кинетической энергии множества молекул. Кристаллическая решётка обладает симметрией, которой не обладают образующие её атомы по отдельности. Во всех этих случаях мы наблюдаем появление новых свойств, которые реальны и измеримы, но которые невозможно обнаружить при изучении отдельных компонентов. Это не мистика и не отрицание физических законов; это проявление того, как организация и взаимодействие создают качества, отсутствующие в элементах.
В нейробиологии стресса эмерджентность проявляется с особой наглядностью при рассмотрении нейронных сетей и их функций. Отдельный нейрон представляет собой клетку, способную генерировать электрические импульсы и высвобождать нейромедиаторы. Он не мыслит, не чувствует, не помнит, не принимает решений. Все эти когнитивные и эмоциональные функции являются эмерджентными свойствами сетей, состоящих из миллиардов нейронов, связанных триллионами синапсов. Паттерн синхронизированной активности миллионов нейронов, измеряемый методами электроэнцефалографии как ритмическая волна определённой частоты, не существует на уровне отдельных клеток; он возникает из их координированного взаимодействия. Состояние сознания, например переход от бодрствования ко сну, представляет собой глобальное изменение режима работы мозга, которое невозможно локализовать в каком-либо одном нейроне или даже в одной структуре; это свойство целостной системы.
Психологические феномены стресса представляют собой, возможно, наиболее драматические примеры эмерджентности. Руминация, характеризующаяся навязчивым прокручиванием негативных мыслей и неспособностью отвлечься от них, является клинически значимым предиктором депрессии и других стресс-связанных расстройств. Этот феномен невозможно редуцировать к активности отдельных нейронов или даже отдельных мозговых структур. Руминация возникает как паттерн динамического взаимодействия между сетью пассивного режима работы мозга, отвечающей за самореферентное мышление, исполнительной сетью, обеспечивающей когнитивный контроль, и эмоциональными системами, включающими миндалину и связанные с ней структуры. Характерная для руминации «застреваемость» в негативном содержании отражает нарушение динамического баланса между этими сетями, их неспособность гибко переключаться между режимами функционирования. Это системное свойство, эмерджентное относительно составляющих его нейронных популяций.
Философская дискуссия о природе эмерджентности традиционно различает два её типа, обозначаемые как «слабая» и «сильная» эмерджентность, и это различие имеет существенные эпистемологические и методологические импликации. Слабая эмерджентность означает, что свойства высокого уровня в принципе вычислимы из полного описания низкого уровня, но это вычисление практически недоступно из-за сложности системы и огромного числа взаимодействий. Поведение погоды, например, слабо эмерджентно относительно молекулярной физики: законы движения молекул воздуха полностью определяют погодные явления, но практическое предсказание погоды требует иных, макроскопических моделей. Сильная эмерджентность предполагает принципиальную невыводимость свойств высокого уровня из законов низкого уровня, даже при полном знании последних. Вопрос о том, является ли сознание и субъективный опыт сильно эмерджентными относительно нейронной активности, остаётся одной из центральных проблем философии сознания и не имеет консенсусного решения.
Для практических целей науки о стрессе вопрос о типе эмерджентности менее важен, чем признание самого факта её существования и методологических следствий из этого признания. Независимо от того, является ли психологическое переживание тревоги в принципе выводимым из полного нейробиологического описания или нет, практически такая дедукция недоступна, и психологический уровень анализа сохраняет свою объяснительную ценность. Это означает, что изучение стресса должно вестись на нескольких уровнях одновременно, с признанием относительной автономии каждого уровня. Психологические закономерности копинга и эмоциональной регуляции, рассматриваемые в пятом модуле курса, не являются просто сокращённым описанием нейробиологических процессов; они представляют собой объяснения на своём уровне, схватывающие паттерны, не видимые с биологической точки зрения. Аналогично, социальные закономерности, рассматриваемые в десятом модуле, обладают своей объяснительной силой, не редуцируемой к психологии индивидов.
Концепция эмерджентности имеет прямые следствия для понимания индивидуальных различий в реакции на стресс. Два человека с идентичными генотипами по всем известным генам уязвимости к стрессу могут радикально различаться по своей фактической уязвимости в силу различий в организации их нейронных сетей, сформировавшихся под влиянием уникальной истории развития. Эта организация является эмерджентным свойством взаимодействия генов и среды на протяжении онтогенеза, и она не предсказуема из знания отдельных генетических вариантов. Более того, два человека с похожей нейробиологией могут по-разному реагировать на стресс в силу различий в когнитивной оценке, которая, в свою очередь, является эмерджентным продуктом их жизненного опыта, убеждений, ценностей, социального контекста. Эта многоуровневая детерминация делает предсказание индивидуальной реакции на стресс принципиально сложной задачей, не решаемой редукцией к какому-либо одному уровню.
Признание эмерджентности не означает отказа от поиска связей между уровнями; напротив, оно направляет этот поиск в продуктивное русло. Задача состоит не в том, чтобы редуцировать высокие уровни к низким, а в том, чтобы понять, как свойства высоких уровней возникают из организации низких уровней. Это требует изучения механизмов связывания уровней, таких как нисходящая причинность, которой посвящён следующий раздел данной темы. В контексте курса это означает, что после изучения биологических механизмов стресса в модулях со второго по четвёртый студенты не должны считать психологическое содержание пятого модуля просто «прикладным» или «описательным». Психологические процессы когнитивной оценки, эмоциональной регуляции, копинга представляют собой реальные явления со своими закономерностями, которые взаимодействуют с биологическими механизмами, но не сводятся к ним. Интегративное понимание стресса требует способности видеть обе стороны этой связи и понимать их взаимную обусловленность.
1.4 Нисходящая причинность: влияние высших уровней организации на низшие
Традиционное редукционистское мышление предполагает исключительно восходящую направленность причинных связей в иерархии уровней организации: молекулярные события определяют клеточные процессы, клеточные процессы определяют функции тканей и органов, органные функции определяют системную физиологию, а физиология в конечном счёте порождает психологические явления и поведение. Эта модель однонаправленной детерминации, при всей её интуитивной привлекательности, оказывается неадекватной для описания реальных биологических систем, в которых причинные влияния распространяются не только снизу вверх, но и сверху вниз. Концепция нисходящей причинности фиксирует этот фундаментальный факт: состояния и процессы на высших уровнях организации способны оказывать каузальное воздействие на процессы низших уровней, модифицируя их протекание способами, которые невозможно предсказать из рассмотрения низшего уровня в изоляции. В науке о стрессе нисходящая причинность проявляется с особой очевидностью, поскольку психологические процессы оценки, интерпретации и произвольной регуляции демонстрируют способность изменять нейробиологические и даже молекулярные параметры стрессового ответа.
Наиболее убедительные демонстрации нисходящей причинности в контексте стресса связаны с феноменом когнитивной переоценки и его измеримыми биологическими последствиями. Когда человек сознательно изменяет интерпретацию потенциально угрожающей ситуации, рассматривая её не как опасность, а как вызов или возможность для роста, это когнитивное изменение, происходящее на психологическом уровне, вызывает каскад измеримых изменений на биологических уровнях. Нейровизуализационные исследования демонстрируют, что успешная когнитивная переоценка сопровождается снижением активности миндалевидного тела и усилением активности префронтальной коры, причём степень подавления миндалины коррелирует с субъективно оцениваемой эффективностью регуляции эмоций. На эндокринном уровне когнитивная переоценка ассоциируется с ослаблением кортизолового ответа на стрессор. На иммунном уровне обнаруживается снижение провоспалительной активации. Все эти изменения инициируются событием психологического порядка и распространяются вниз по уровням организации, что представляет собой парадигматический случай нисходящей причинности.
Феномен плацебо представляет собой, пожалуй, наиболее драматическую и хорошо документированную демонстрацию нисходящей причинности в медицине и науке о стрессе. Когда пациент получает инертное вещество, но верит в его терапевтическую эффективность, это убеждение, существующее на когнитивном уровне, способно вызывать реальные физиологические изменения, включая высвобождение эндогенных опиоидов, модуляцию дофаминергической нейротрансмиссии, изменения иммунной функции и даже экспрессии генов. Плацебо-анальгезия, например, опосредуется активацией тех же нейрохимических систем, которые активируются реальными анальгетиками, и может быть заблокирована антагонистом опиоидных рецепторов налоксоном. Это означает, что ожидание облегчения боли, являющееся психологическим состоянием, причинно вызывает высвобождение конкретных молекул в конкретных нейронных цепях. Редукционистская модель, допускающая только восходящую причинность, не способна объяснить этот феномен; его объяснение требует признания реальности нисходящих каузальных влияний.
Философская проблематичность нисходящей причинности связана с вопросом о том, как ментальные состояния, традиционно понимаемые как нематериальные, могут оказывать каузальное воздействие на материальные процессы в мозге и теле. Этот вопрос уходит корнями в картезианский дуализм, постулировавший принципиальное различие между мыслящей субстанцией и протяжённой субстанцией и оставивший неразрешённой проблему их взаимодействия. Современная нейронаука преодолевает этот дуализм через признание того, что ментальные состояния не являются нематериальными сущностями, отдельными от мозга; они представляют собой паттерны нейронной активности, организованные на высоком уровне. Мысль или убеждение есть определённая конфигурация активности распределённых нейронных сетей, и эта конфигурация, как целостный паттерн, способна оказывать регулирующее влияние на активность составляющих её элементов и связанных с ними систем. Нисходящая причинность в этом понимании не требует мистического воздействия нематериального на материальное; она представляет собой влияние паттерна организации на динамику компонентов.
Механизмы реализации нисходящей причинности в нервной системе включают множество нейробиологических путей, через которые высокоуровневые когнитивные процессы модулируют низкоуровневую нейронную и физиологическую активность. Префронтальная кора, являющаяся субстратом высших когнитивных функций, включая произвольную регуляцию поведения и эмоций, посылает обширные нисходящие проекции к подкорковым структурам, включая миндалину, гипоталамус, стволовые центры вегетативной регуляции. Эти проекции позволяют когнитивным процессам, реализуемым в коре, модулировать активность эволюционно более древних систем, ответственных за автоматические стрессовые реакции. Нейромодуляторные системы, включая норадреналиновую, серотониновую, дофаминовую, служат дополнительными каналами нисходящего влияния, позволяя глобальным состояниям мозга регулировать чувствительность локальных нейронных сетей. Вегетативная нервная система транслирует нисходящие влияния центральной нервной системы на периферические органы, а гипоталамо-гипофизарные оси обеспечивают нисходящий контроль над эндокринными железами.
Эпигенетические механизмы представляют особенно интригующий канал нисходящей причинности, через который психологический опыт может оказывать долговременное влияние на экспрессию генов. Исследования показывают, что психотерапевтические интервенции, представляющие собой воздействия на когнитивном и эмоциональном уровнях, способны вызывать измеримые изменения в эпигенетических метках, регулирующих активность генов, связанных со стрессовым ответом. Медитативные практики ассоциируются с изменениями в метилировании генов, связанных с воспалением и кортизоловой регуляцией. Когнитивно-поведенческая терапия посттравматического стрессового расстройства сопровождается нормализацией эпигенетических изменений, индуцированных травмой. Эти находки демонстрируют, что влияние психологических интервенций проникает до молекулярного уровня, изменяя то, как генетическая информация читается клетками. Нисходящая причинность здесь проявляется в буквальном смысле: изменение на уровне сознательного опыта и поведения вызывает изменение на уровне химических модификаций ДНК.
Концепция нисходящей причинности имеет существенные импликации для понимания терапевтических интервенций при стресс-связанных расстройствах, и эти импликации непосредственно связаны с содержанием девятого модуля курса, посвящённого интервенциям. Традиционное противопоставление фармакотерапии как «биологического» лечения и психотерапии как «психологического» лечения оказывается поверхностным в свете понимания нисходящей причинности. Психотерапия является биологическим вмешательством в том смысле, что она изменяет мозг через нисходящие каузальные пути; нейровизуализационные исследования убедительно демонстрируют структурные и функциональные изменения мозга после успешной психотерапии. Фармакотерапия, в свою очередь, оказывает психологические эффекты, изменяя субъективный опыт, когнитивные процессы, эмоциональную регуляцию. Оба типа интервенций работают через биопсихологическую систему, в которой причинные влияния распространяются в обоих направлениях. Это понимание открывает возможности для интеграции и потенцирования различных терапевтических модальностей.
Признание нисходящей причинности также трансформирует понимание роли субъективного опыта и произвольного контроля в регуляции стресса. Если бы причинность была исключительно восходящей, произвольные усилия по управлению стрессом были бы эпифеноменальными, не имеющими реального влияния на биологические процессы. Человек был бы пассивным наблюдателем своих стрессовых реакций, определяемых молекулярными событиями вне его контроля. Нисходящая причинность обосновывает реальную эффективность произвольных стратегий совладания: когнитивная переоценка, направленное внимание, регуляция дыхания, осознанное расслабление мышц — все эти техники, рассматриваемые в модуле об интервенциях, работают именно потому, что психологические и поведенческие изменения причинно влияют на физиологические процессы. Это не самовнушение и не иллюзия контроля; это реализация нисходящей причинности через конкретные нейробиологические механизмы.
Вместе с тем концепция нисходящей причинности не должна интерпретироваться как отрицание важности биологических оснований или как утверждение неограниченной власти сознания над телом. Нисходящие влияния реализуются через материальные механизмы и ограничены структурой этих механизмов. Человек не может произвольно остановить секрецию кортизола одним усилием воли; он может только инициировать когнитивные и поведенческие процессы, которые через опосредующие нейронные и эндокринные пути постепенно модулируют стрессовый ответ. При некоторых состояниях, когда биологические механизмы серьёзно нарушены, возможности нисходящей регуляции могут быть ограничены, и требуется прямое фармакологическое вмешательство для восстановления субстрата, через который могла бы осуществляться произвольная регуляция. Адекватное понимание нисходящей причинности предполагает признание как её реальности, так и её ограничений, что соответствует интегративному подходу, развиваемому на протяжении всего курса.
1.5 Множественная реализуемость: различные механизмы порождения единого феномена
Концепция множественной реализуемости представляет собой одно из ключевых понятий философии сознания и философии науки, имеющее глубокие импликации для понимания природы стресса и стресс-связанных расстройств. Суть этой концепции состоит в признании того, что один и тот же феномен на высшем уровне описания может быть реализован через различные механизмы на низшем уровне. Иными словами, функциональное или феноменологическое тождество не предполагает структурного тождества: разные физические системы могут выполнять одну и ту же функцию, разные нейробиологические паттерны могут порождать одно и то же психологическое состояние, разные молекулярные пути могут приводить к одному и тому же клиническому симптому. Это простое на первый взгляд наблюдение имеет далеко идущие последствия для исследовательских стратегий, диагностических подходов и терапевтических вмешательств в области стресса, поскольку оно ставит под вопрос поиск единственного «правильного» механизма для каждого явления.
В контексте науки о стрессе множественная реализуемость наиболее очевидно проявляется в феноменологии усталости — одного из наиболее распространённых симптомов хронического стресса и стресс-связанных расстройств. Субъективное переживание усталости, истощённости, отсутствия энергии может возникать через принципиально различные биологические механизмы. У одного пациента усталость может быть следствием хронического воспаления: провоспалительные цитокины, такие как интерлейкин-6 и фактор некроза опухоли альфа, воздействуют на мозг и индуцируют так называемое «болезненное поведение», включающее усталость как центральный компонент. У другого пациента усталость может быть результатом митохондриальной дисфункции: энергетические станции клеток работают неэффективно, производят недостаточно аденозинтрифосфата, и организм испытывает буквальный дефицит клеточной энергии. У третьего пациента усталость может быть связана с нарушениями циркадной регуляции: десинхронизация внутренних биологических часов нарушает нормальные циклы активности и отдыха. У четвёртого механизм может быть преимущественно дофаминергическим: снижение мотивационного драйва при нарушении мезолимбической дофаминовой системы переживается как отсутствие энергии и инициативы.
Феномен депрессии представляет собой, возможно, наиболее клинически значимый пример множественной реализуемости в психиатрии стресс-связанных расстройств. Диагностическая категория большого депрессивного расстройства объединяет пациентов с общим набором симптомов, но исследования последних десятилетий убедительно продемонстрировали глубокую биологическую гетерогенность этой популяции. Выделяются подтипы депрессии с преобладанием воспалительного компонента, характеризующиеся повышенными маркерами воспаления и лучшим ответом на противовоспалительные интервенции. Идентифицируются подтипы с преимущественной гиперактивностью гипоталамо-гипофизарно-надпочечниковой оси, проявляющиеся гиперкортизолемией и специфическими нейроэндокринными нарушениями. Описываются подтипы с акцентом на нарушениях нейропластичности, где центральную роль играет дефицит нейротрофического фактора мозга и нарушение синаптической пластичности в ключевых нейронных цепях. Каждый из этих подтипов представляет собой различную биологическую реализацию того, что на клиническом уровне описывается как «депрессия».
Множественная реализуемость имеет критические импликации для поиска биомаркеров стресса и стресс-связанных расстройств, что непосредственно связано с содержанием шестого модуля курса, посвящённого измерению. Если один и тот же клинический феномен может реализовываться через разные биологические механизмы, то поиск единственного универсального биомаркера этого феномена обречён на неудачу. Повышенный уровень кортизола является биомаркером для одних подтипов стресс-связанных расстройств, но не для других; более того, при некоторых хронических состояниях наблюдается парадоксальная гипокортизолемия. Воспалительные маркеры, такие как С-реактивный белок, информативны для подгруппы пациентов с воспалительным профилем, но неспецифичны для депрессии или посттравматического расстройства в целом. Это объясняет, почему десятилетия поисков диагностических биомаркеров для психических расстройств не привели к их внедрению в клиническую практику: биомаркер, который хорошо работает для подгруппы, размывается и теряет чувствительность при применении к гетерогенной популяции.
Терапевтические следствия множественной реализуемости состоят в необходимости дифференцированного подхода к лечению, который учитывает конкретный механизм, лежащий в основе симптоматики у данного пациента. Антидепрессанты, действующие через серотониновую систему, могут быть высокоэффективны для пациентов, у которых патофизиология депрессии действительно связана с серотонинергической дисфункцией, но неэффективны или малоэффективны для пациентов с преимущественно воспалительным или метаболическим механизмом. Это объясняет хорошо известный клинический факт: только около трети пациентов с депрессией достигают ремиссии на первом назначенном антидепрессанте, и значительная часть пациентов остаётся резистентной к стандартной фармакотерапии. Концепция множественной реализуемости указывает на то, что это не провал лечения, а неадекватность подхода, предполагающего единый механизм для всех случаев. Стратификация пациентов по биологическим подтипам и подбор терапии, нацеленной на конкретный механизм, представляет собой перспективное направление развития персонализированной медицины.
Генетические исследования стресс-связанных расстройств наглядно демонстрируют множественную реализуемость на молекулярном уровне. Полногеномные ассоциативные исследования идентифицировали сотни генетических вариантов, каждый из которых вносит малый вклад в риск развития депрессии, тревожных расстройств или посттравматического стрессового расстройства. Эти варианты затрагивают гены, вовлечённые в самые разные биологические процессы: нейротрансмиссию, воспаление, нейропластичность, метаболизм, циркадные ритмы, функцию гипоталамо-гипофизарно-надпочечниковой оси. Разные индивиды могут иметь повышенный генетический риск через разные комбинации вариантов, затрагивающих разные биологические пути, но конвергировать к сходному фенотипу уязвимости к стрессу. Эта генетическая гетерогенность является молекулярным отражением множественной реализуемости: множество генетических путей ведут к тому, что на фенотипическом уровне выглядит как единое состояние.
Концепция эквифинальности, заимствованная из общей теории систем, дополняет понимание множественной реализуемости динамическим измерением. Эквифинальность означает, что открытая система может достигать одного и того же конечного состояния из различных начальных условий и различными путями. В контексте развития стресс-связанных расстройств это проявляется в том, что сходная клиническая картина может быть результатом различных траекторий развития. Один человек может прийти к депрессии через раннюю травму, эпигенетическое программирование систем стресса, формирование уязвимых паттернов привязанности и когнитивных схем. Другой может достичь сходного состояния через хронический профессиональный стресс во взрослом возрасте, истощение адаптационных ресурсов, накопление аллостатической нагрузки. Третий путь может включать соматическое заболевание, хроническое воспаление, вторичные нейропсихиатрические эффекты. Все три траектории конвергируют к тому, что диагностически классифицируется как депрессия, но представляют собой различные этиопатогенетические сущности.
Множественная реализуемость ставит под вопрос традиционную нозологию психических расстройств, основанную на синдромальных описаниях, и открывает путь к трансдиагностическим и механистическим подходам, которые рассматриваются в седьмом модуле курса. Если за одним диагнозом скрываются различные биологические реальности, то категориальная диагностика имеет ограниченную биологическую валидность. Альтернативный подход состоит в идентификации базовых патофизиологических механизмов или процессов, которые могут присутствовать при различных расстройствах и отсутствовать у части пациентов с одним и тем же диагнозом. Воспалительный процесс, например, может быть трансдиагностическим механизмом, присутствующим при депрессии, тревоге, посттравматическом расстройстве, шизофрении, но характерным не для всех пациентов с каждым из этих диагнозов. Терапия, нацеленная на этот механизм, потенциально эффективна для пациентов с воспалительным профилем независимо от их категориального диагноза.
Философское значение множественной реализуемости выходит за рамки практических приложений и затрагивает фундаментальные вопросы о природе психологических и клинических категорий. Если депрессия множественно реализуема, является ли она единой сущностью или конгломератом различных состояний, искусственно объединённых общим названием? Этот вопрос не имеет однозначного ответа и связан с более широкими дебатами о природе естественных видов в науке. Прагматический подход, принятый в данном курсе, состоит в признании полезности клинических категорий для коммуникации и организации знания при одновременном понимании их биологической гетерогенности. Это означает, что, говоря о «стрессе» или «депрессии», мы используем полезные обобщения, которые, однако, не должны затемнять лежащее за ними многообразие механизмов и траекторий развития.
1.6 Интегративный плюрализм: необходимость многоуровневых объяснений
Рассмотрение редукционизма, эмерджентности, нисходящей причинности и множественной реализуемости логически приводит к позиции интегративного плюрализма как наиболее адекватной эпистемологической рамки для науки о стрессе. Интегративный плюрализм представляет собой философскую позицию, согласно которой полное понимание сложного феномена требует объяснений на множественных уровнях анализа, причём эти объяснения не сводимы друг к другу, но и не изолированы, а находятся в отношениях взаимного дополнения и констрейнинга. Применительно к стрессу это означает, что молекулярные, клеточные, системные, психологические и социальные объяснения не являются конкурирующими альтернативами, из которых нужно выбрать «правильную», и не являются независимыми описаниями, не связанными друг с другом. Они представляют собой различные, но взаимосвязанные перспективы на единый многоуровневый феномен, и задача интегративной науки состоит в том, чтобы связать эти перспективы в когерентную картину.
Необходимость плюралистического подхода вытекает из самой природы стресса как феномена, конституированного на множественных уровнях организации одновременно. Стресс не является ни чисто биологическим событием, происходящим в теле и лишь отражающимся в сознании, ни чисто психологическим переживанием, которое вторично влияет на тело. Стресс с самого начала является биопсихосоциальным: угроза воспринимается и оценивается психологически, эта оценка немедленно транслируется в нейробиологические паттерны, которые запускают каскады гормональных и иммунных изменений, а весь этот процесс разворачивается в социальном контексте, который определяет природу угроз, доступные ресурсы совладания и смыслы, приписываемые переживаниям. Попытка объяснить стресс исключительно на одном уровне, игнорируя другие, неизбежно упускает существенные аспекты феномена. Молекулярная редукция теряет смыслы и контексты; социологический анализ теряет механизмы и индивидуальные различия; психологическое описание теряет биологические ограничения и возможности.
Отношения между уровнями объяснения в интегративном плюрализме не являются ни иерархическими в смысле подчинения высших уровней низшим, ни произвольно-эклектическими в смысле независимого сосуществования. Они представляют собой отношения взаимного констрейнинга: объяснение на одном уровне ограничивает возможные объяснения на других уровнях и должно быть совместимым с ними. Психологическая теория копинга, постулирующая определённые когнитивные стратегии регуляции стресса, должна быть совместима с нейробиологическими данными о механизмах эмоциональной регуляции; если нейробиология не обнаруживает субстрата для постулируемых процессов, психологическая теория требует пересмотра. Аналогично, нейробиологическая модель должна быть совместима с психологическими и клиническими наблюдениями; если модель предсказывает эффекты, которые не наблюдаются на уровне поведения или переживания, модель неадекватна. Это взаимное ограничение создаёт связность между уровнями и предотвращает как редукционистскую элиминацию, так и беспринципный эклектизм.
Практическая реализация интегративного плюрализма в науке о стрессе требует преодоления дисциплинарных барьеров, традиционно разделяющих молекулярную биологию, нейронауки, психологию, социологию и клинические дисциплины. Каждая из этих областей развила собственный концептуальный аппарат, методологию, стандарты доказательности, профессиональные сообщества и журналы. Исследователь, работающий на молекулярном уровне, может не читать психологическую литературу и не понимать её концепции; клинический психолог может не следить за достижениями нейроэндокринологии. Эта дисциплинарная специализация имеет свои основания в необходимости глубокой экспертизы, но она создаёт препятствия для интеграции знания. Преодоление этих барьеров требует институциональных изменений: междисциплинарных исследовательских программ, журналов, принимающих многоуровневые исследования, образовательных программ, формирующих интегративное мышление. Данный курс представляет собой попытку такого образовательного решения.
Интегративный подход к пониманию стресса имеет прямые следствия для терапевтической практики. Если стресс конституирован на множественных уровнях, то и терапевтическое воздействие должно учитывать множественные уровни. Фармакотерапия воздействует преимущественно на молекулярный и клеточный уровни, модифицируя нейротрансмиссию, рецепторную чувствительность, экспрессию генов. Психотерапия работает на психологическом уровне с когнициями, эмоциями, поведенческими паттернами, но через нисходящую причинность влияет и на биологию. Социальные интервенции изменяют контекст, в котором разворачивается стресс: условия труда, качество отношений, доступ к ресурсам. Оптимальная стратегия помощи может комбинировать вмешательства на разных уровнях, используя их синергию. Для одного пациента ключевым может быть фармакологическая стабилизация, создающая условия для психотерапевтической работы; для другого психотерапия может быть достаточна сама по себе; для третьего критическим является изменение социальной ситуации, без которого никакая индивидуальная терапия не будет эффективной.
Интегративный плюрализм не следует смешивать с беспринципным эклектизмом или релятивизмом, утверждающим равную ценность любых объяснений. Плюрализм признаёт множественность легитимных перспектив, но это не означает, что все перспективы равно адекватны для любой задачи. Для понимания молекулярных механизмов действия лекарственного препарата биохимический анализ более релевантен, чем социологический. Для понимания социального неравенства в распространённости стресс-связанных расстройств социологический анализ более информативен, чем молекулярный. Для понимания индивидуального опыта переживания стресса феноменологический и психологический подходы могут быть более адекватны, чем нейровизуализация. Выбор уровня анализа определяется исследовательским вопросом и практической задачей. Интегративный плюрализм настаивает на том, что ни один уровень не имеет привилегированного статуса единственно правильного, но это не исключает, а предполагает обоснованный выбор уровня для конкретной цели.
Эпистемологическое обоснование интегративного плюрализма связано с признанием того, что научные объяснения служат различным целям и отвечают на различные типы вопросов. Вопрос «как?», запрашивающий механизм, требует иного типа объяснения, чем вопрос «почему?», запрашивающий функцию или причину, и оба они отличаются от вопроса «что это значит?», запрашивающего смысл и интерпретацию. Молекулярное объяснение превосходно отвечает на определённые «как»-вопросы о механизмах сигнальной трансдукции, но не отвечает на вопросы о смысле переживания стресса для человека или о социальных условиях, порождающих хронический стресс в определённых популяциях. Для полного понимания феномена стресса нужны ответы на все эти вопросы, и они приходят с разных уровней анализа. Плюрализм здесь не слабость, а адекватный ответ на многомерность реальности.
Структура данного курса воплощает принципы интегративного плюрализма в организации учебного материала. Движение от модуля к модулю представляет собой последовательное рассмотрение различных уровней анализа стресса: от молекулярного и клеточного в первом модуле, через нейробиологический во втором, нейроэндокринный в третьем, иммунный и метаболический в четвёртом, к психологическому в пятом, методологическому в шестом, патологическому в седьмом и восьмом, терапевтическому в девятом, контекстуальному в десятом и перспективному в одиннадцатом. Эта последовательность не означает иерархии важности; она отражает логику постепенного выстраивания многоуровневого понимания. Сквозные связи между модулями, обозначенные в структуре курса, указывают на интеграцию уровней: то, как молекулярные механизмы из третьего модуля связаны с психологическими процессами из пятого, как измерения из шестого модуля применяются к патологии из седьмого, как понимание механизмов из первых модулей обосновывает интервенции из девятого.
Формирование интегративного мышления у студентов является одной из центральных образовательных целей курса. Это означает развитие способности видеть феномен стресса одновременно с нескольких точек зрения, понимать связи между уровнями, выбирать адекватный уровень анализа для конкретной задачи, интегрировать информацию из разных источников в когерентное понимание. Такое мышление не формируется автоматически при последовательном изложении материала разных уровней; оно требует целенаправленных усилий по демонстрации связей, по анализу случаев через призму разных уровней, по критическому рассмотрению ограничений каждой перспективы. Настоящий урок, посвящённый философским основаниям многоуровневого анализа, создаёт концептуальную рамку для такой интеграции. Последующие модули наполняют эту рамку конкретным содержанием, а заключительный модуль рассматривает перспективы развития интегративных подходов в науке о стрессе.
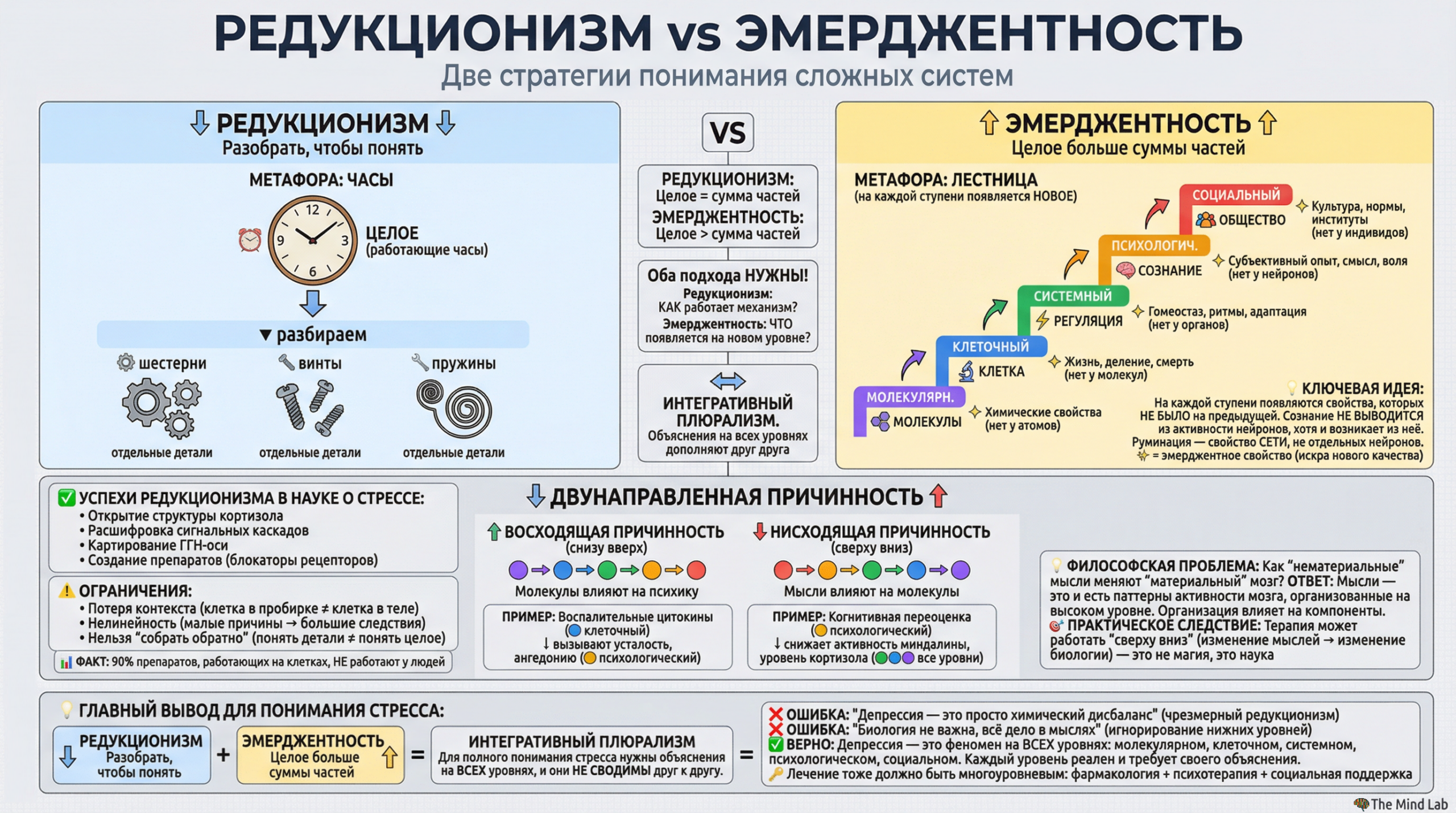
2. Многоуровневое моделирование: методологические вызовы исследования стресса
2.1 Концептуальная проблема: определение границ уровней
Переход от философского признания многоуровневой природы стресса к практическому исследованию этого феномена немедленно сталкивается с фундаментальной концептуальной трудностью, которая состоит в определении того, где заканчивается один уровень анализа и начинается другой. На первый взгляд разделение на молекулярный, клеточный, системный, психологический и социальный уровни представляется интуитивно ясным и самоочевидным, однако при более внимательном рассмотрении границы между этими уровнями оказываются размытыми, подвижными и в значительной мере зависящими от перспективы исследователя. Эта проблема не является лишь философским затруднением, не имеющим практических последствий; напротив, она непосредственно влияет на планирование исследований, выбор методов, интерпретацию результатов и возможности интеграции данных, полученных в разных исследовательских традициях. Осознание условности и аналитического характера границ между уровнями составляет необходимое условие для грамотного многоуровневого исследования и предохраняет от наивного реализма, полагающего, что природа сама разделена на дискретные уровни, которые исследователь лишь обнаруживает.
Рассмотрим конкретный пример, демонстрирующий размытость границ между уровнями: цитокины, такие как интерлейкин-6 или фактор некроза опухоли альфа, играющие центральную роль в воспалительных процессах при хроническом стрессе. С одной стороны, цитокины представляют собой белковые молекулы с определённой химической структурой, аминокислотной последовательностью, трёхмерной конфигурацией, и в этом смысле они принадлежат молекулярному уровню анализа. С другой стороны, цитокины производятся конкретными клетками, преимущественно иммунными, в ответ на определённые сигналы, и их понимание невозможно без рассмотрения клеточного контекста их синтеза и секреции. Более того, цитокины функционируют как сигнальные молекулы, опосредующие коммуникацию между клетками и между системами организма, включая взаимодействие иммунной системы с нервной и эндокринной, что помещает их также на системный уровень анализа. Наконец, эффекты цитокинов на мозг проявляются в изменениях настроения, когнитивных функций, мотивации, что связывает их с психологическим уровнем. Вопрос о том, к какому уровню «на самом деле» принадлежат цитокины, не имеет однозначного ответа, поскольку сам вопрос некорректен: цитокины участвуют в процессах на всех этих уровнях одновременно.
Аналогичная неопределённость обнаруживается при рассмотрении нейроанатомических структур, традиционно относимых к системному уровню анализа. Миндалевидное тело, или амигдала, обычно описывается как компонент лимбической системы, ответственный за обработку эмоционально значимых стимулов и формирование страха. В таком описании миндалина выступает как системная единица, взаимодействующая с другими системными единицами, такими как гиппокамп, префронтальная кора, гипоталамус. Однако при более детальном рассмотрении миндалина оказывается сложной структурой, состоящей из множества ядер с различными функциями, различными входами и выходами, различными популяциями нейронов с различными нейрохимическими характеристиками. Каждое ядро может изучаться как самостоятельная единица, и уровень анализа смещается от системного к более локальному. Далее, каждое ядро состоит из нейронов, и понимание функции миндалины требует понимания электрофизиологических свойств этих нейронов, их синаптических связей, рецепторного состава, что переводит анализ на клеточный уровень. Наконец, функция нейрона определяется молекулярными процессами экспрессии генов, синтеза белков, ионного транспорта, что возвращает нас к молекулярному уровню. Миндалина, таким образом, не принадлежит исключительно одному уровню; она представляет собой объект, который может быть описан на разных уровнях в зависимости от исследовательского вопроса.
Проблема определения границ уровней усугубляется тем, что различные феномены имеют различные характерные пространственные и временные масштабы, которые не всегда соответствуют традиционным уровням анализа. Пространственные масштабы в науке о стрессе охватывают диапазон от нанометров, на которых происходят конформационные изменения рецепторных белков, до километров и континентов, на которых рассматриваются популяционные различия в распространённости стресс-связанных расстройств. Между этими крайностями располагаются масштабы синапсов, нейронов, нейронных ансамблей, мозговых структур, целого мозга, организма, социальных групп, сообществ, обществ. Временные масштабы простираются от миллисекунд, за которые происходит синаптическая передача, до десятилетий, на протяжении которых накапливается аллостатическая нагрузка и разворачиваются последствия ранней травмы. Процессы на разных временных масштабах взаимодействуют: миллисекундные события синаптической пластичности, повторяясь, приводят к минутным изменениям синаптической силы, которые, консолидируясь, создают долговременные структурные изменения, формирующие память и поведенческие паттерны на протяжении лет. Традиционные уровни анализа лишь приблизительно соответствуют определённым пространственно-временным масштабам и не исчерпывают их многообразия.
Осознание условности границ между уровнями приводит к пониманию того, что деление на уровни представляет собой аналитическую конструкцию, создаваемую исследователем для организации знания и планирования исследований, а не открытие естественных разломов в структуре реальности. Природа не разделена на молекулярный, клеточный и системный уровни; эти категории являются концептуальными инструментами, которые мы накладываем на непрерывную ткань биологических процессов для их понимания и изучения. Признание конструктивного характера уровней не обесценивает их; напротив, оно позволяет использовать их более осознанно и гибко. Исследователь, понимающий условность уровней, может свободнее переключаться между ними, выбирая наиболее адекватный уровень для своего вопроса, и легче интегрировать данные с разных уровней, не будучи связанным ригидными дисциплинарными границами. Эта эпистемологическая рефлексия особенно важна для междисциплинарных исследований стресса, где встречаются традиции с различными представлениями о релевантных уровнях анализа.
Выбор уровней анализа в конкретном исследовании должен определяться исследовательским вопросом, а не традицией дисциплины или доступностью методов, хотя последние факторы неизбежно влияют на практику. Если исследователь спрашивает, как хронический стресс влияет на экспрессию генов, связанных с воспалением, он выбирает молекулярный уровень анализа с возможным расширением на клеточный для понимания того, в каких клетках происходят эти изменения. Если вопрос касается того, как организационные факторы рабочей среды связаны с выгоранием сотрудников, релевантны социальный и психологический уровни. Если же исследователь стремится понять, как социальные факторы «проникают под кожу» и изменяют биологию, ему необходимо связать социальный, психологический и биологический уровни в едином исследовательском дизайне. Ключевым здесь является осознанность выбора: исследователь должен понимать, почему он выбирает определённые уровни, что этот выбор позволяет увидеть и что оставляет за пределами рассмотрения. Эта рефлексивность предохраняет от неосознанного сужения перспективы и от необоснованных обобщений за пределы изученных уровней.
Практические следствия проблемы границ уровней для организации данного курса состоят в том, что разделение материала на модули, посвящённые различным уровням анализа, является дидактической условностью, а не отражением реальных разрывов в предмете изучения. Первый модуль, посвящённый организму как адаптивной системе, начинает с клеточного уровня и поднимается к системной интеграции. Второй модуль фокусируется на нервной системе и мозге. Третий рассматривает нейроэндокринные оси. Четвёртый обращается к иммунитету, воспалению и метаболизму. Эти разделения удобны для последовательного изложения, но не должны создавать впечатление, что нервная, эндокринная и иммунная системы функционируют раздельно; напротив, их интеграция подчёркивается на протяжении всего курса. Пятый модуль о психологии стресса не противопоставляется биологическим модулям, а показывает психологический уровень как эмерджентный относительно биологии, но имеющий собственные закономерности. Сквозные связи между модулями, отмеченные в структуре курса, призваны восстановить ту целостность, которая неизбежно нарушается при последовательном изложении.
Методологическое значение обсуждаемой проблемы состоит также в необходимости осторожности при переносе результатов между уровнями. Закономерность, установленная на одном уровне анализа, не автоматически переносится на другие уровни. Если показано, что на молекулярном уровне кортизол подавляет определённые иммунные функции, это не означает автоматически, что люди с высоким уровнем кортизола будут иметь ослабленный иммунитет, поскольку на системном уровне действуют компенсаторные механизмы, контекстуальные факторы, индивидуальные различия. Если на психологическом уровне показано, что социальная поддержка снижает воспринимаемый стресс, это не означает автоматически, что она снижает кортизол или воспаление, хотя такие связи возможны и эмпирически изучаются. Каждый переход между уровнями требует отдельного эмпирического обоснования и теоретического объяснения механизмов связи. Это предостережение особенно важно для трансляционных исследований, стремящихся перенести результаты с фундаментального уровня на клинический, где разрыв между лабораторными находками и терапевтической эффективностью часто связан именно с непредвиденными межуровневыми взаимодействиями.
Философская традиция анализа уровней организации в биологии предлагает несколько подходов к осмыслению обсуждаемой проблемы. Один подход, восходящий к общей теории систем Людвига фон Берталанфи, рассматривает уровни как уровни сложности, где каждый последующий уровень характеризуется большей структурной и функциональной сложностью. Другой подход, развитый в философии биологии, определяет уровни через отношения композиции: клетки составлены из молекул, ткани из клеток, органы из тканей и так далее. Третий подход, более прагматический, определяет уровни через научные дисциплины и их методы: молекулярный уровень изучается биохимией, клеточный — клеточной биологией, психологический — психологией. Каждый из этих подходов имеет свои достоинства и ограничения, и ни один не предлагает однозначного решения проблемы границ. Для целей науки о стрессе наиболее продуктивным представляется прагматический плюрализм, который признаёт множественность возможных способов выделения уровней и выбирает наиболее адекватный в зависимости от исследовательской задачи.
Завершая рассмотрение проблемы границ уровней, необходимо подчеркнуть, что эта проблема не является препятствием для многоуровневого исследования, а скорее условием его правильного понимания и осуществления. Размытость границ не означает, что выделение уровней бесполезно; оно означает, что это выделение должно быть осознанным, гибким и обоснованным применительно к конкретному исследовательскому контексту. Исследователь, работающий на стыке молекулярного и клеточного уровней, не должен тратить усилия на определение того, к какому уровню «на самом деле» принадлежит изучаемый феномен; он должен выбрать концептуальную рамку, наиболее продуктивную для его вопроса, и быть готовым переключаться между рамками по мере необходимости. Эта методологическая гибкость является отличительной чертой зрелого многоуровневого мышления, формирование которого составляет одну из целей данного курса.
2.2 Операционализация переменных на разных уровнях
Если проблема границ уровней касается концептуальной организации многоуровневого исследования, то проблема операционализации относится к переходу от концептуального к эмпирическому, от абстрактных понятий к конкретным измерениям. Операционализация представляет собой процедуру определения абстрактного конструкта через операции его измерения, и в многоуровневом исследовании стресса эта процедура сталкивается с особыми трудностями, связанными с необходимостью измерять концептуально связанные, но эмпирически различные феномены на разных уровнях анализа. Центральный вопрос состоит в следующем: когда мы измеряем кортизол в слюне, воспринимаемый стресс по психологическому опроснику и социально-экономический статус через индикаторы дохода и образования, измеряем ли мы один и тот же конструкт «стресс» на разных уровнях или три разных феномена, лишь связанных между собой? Ответ на этот вопрос имеет критическое значение для интерпретации результатов и для понимания того, что именно мы узнаём о стрессе, используя различные методы измерения.
Принципиальное различие между конструктом и индикатором составляет отправную точку для понимания проблемы операционализации. Конструкт представляет собой теоретическое понятие, обозначающее ненаблюдаемую напрямую сущность или процесс, о существовании которого мы заключаем на основании наблюдаемых проявлений. «Стресс» как конструкт обозначает состояние или процесс, связанный с восприятием угрозы или требования, превышающего доступные ресурсы, и сопровождающийся определёнными биологическими, психологическими и поведенческими изменениями. Индикатор представляет собой конкретную измеримую переменную, которая, как предполагается, отражает или манифестирует конструкт. Концентрация кортизола в слюне является индикатором активности гипоталамо-гипофизарно-надпочечниковой оси, которая, в свою очередь, является компонентом биологического стрессового ответа. Балл по шкале воспринимаемого стресса является индикатором субъективного переживания стресса. Связь между конструктом и индикатором всегда косвенная и опосредованная теоретическими предположениями о том, что индикатор действительно отражает конструкт.
Проблема концептуальной эквивалентности возникает, когда мы пытаемся соотнести индикаторы стресса, измеренные на разных уровнях, и задаёмся вопросом, измеряют ли они «одно и то же». Эмпирические исследования последовательно демонстрируют, что корреляции между индикаторами стресса на разных уровнях умеренные или слабые. Связь между воспринимаемым стрессом, измеренным опросниками, и кортизолом неустойчива и часто незначима; люди с высоким субъективным стрессом не обязательно имеют высокий кортизол, и наоборот. Связь между объективными стрессорами среды и субъективным стрессом также далека от совершенной; одни и те же объективные обстоятельства вызывают различные субъективные реакции у разных людей. Эти слабые корреляции не означают, что измерения ненадёжны или что понятие стресса бессмысленно; они отражают тот факт, что индикаторы на разных уровнях измеряют разные аспекты многомерного феномена, которые связаны между собой, но не тождественны.
Интерпретация слабых межуровневых корреляций требует понимания того, что стресс не является унитарным конструктом, одинаково проявляющимся на всех уровнях, а представляет собой сложный многокомпонентный феномен с частично независимыми измерениями. Биологический стресс-ответ, психологическое переживание стресса и стрессоры среды представляют собой различные, хотя и взаимосвязанные, компоненты. Человек может иметь высокую биологическую реактивность на стрессоры при низком субъективном переживании стресса, если он использует эффективные стратегии эмоциональной регуляции, которые модулируют сознательный опыт, но не полностью блокируют физиологический ответ. Другой человек может субъективно переживать высокий стресс при умеренной биологической активации, если его когнитивные оценки катастрофичны. Третий может находиться в объективно стрессогенной среде, но не воспринимать её как стрессовую в силу привыкания, копинг-ресурсов или особенностей атрибуции. Эти паттерны диссоциации между уровнями являются информативными, а не артефактами измерения; они указывают на сложность механизмов связи между объективной ситуацией, её субъективной оценкой и биологическим ответом.
Методологическое решение проблемы концептуальной эквивалентности состоит не в поиске единственного «правильного» индикатора стресса, а в использовании множественных индикаторов на каждом уровне и в теоретически обоснованном понимании того, что именно каждый индикатор измеряет. На биологическом уровне стрессовый ответ может индексироваться через кортизол, но также через маркеры симпатической активации, такие как катехоламины или вариабельность сердечного ритма, через воспалительные маркеры, такие как С-реактивный белок или интерлейкин-6, через эпигенетические изменения, через показатели аллостатической нагрузки. Каждый из этих индикаторов отражает различные компоненты или аспекты биологического стресс-ответа, и их комбинация даёт более полную картину, чем любой отдельный показатель. На психологическом уровне воспринимаемый стресс может дополняться измерениями тревоги, депрессивности, качества сна, копинг-стратегий, самоэффективности. На социальном уровне индекс жизненных событий может сочетаться с оценкой хронических стрессоров, социальной поддержки, социально-экономического статуса, характеристик рабочей среды.
Латентно-переменное моделирование представляет собой статистический подход, позволяющий формально работать с различием между конструктом и индикатором в многоуровневых исследованиях. В этом подходе конструкт моделируется как латентная, то есть ненаблюдаемая напрямую, переменная, которая объясняет ковариацию между множественными наблюдаемыми индикаторами. Если три разных опросника стресса коррелируют друг с другом, можно постулировать латентный фактор «воспринимаемый стресс», который является общей причиной этих корреляций. Аналогично, латентный фактор «биологическая активация стресса» может объяснять ковариацию между кортизолом, частотой сердечных сокращений и кожно-гальванической реакцией. Структурные модели затем могут оценивать связи между латентными факторами на разных уровнях, очищенные от ошибки измерения отдельных индикаторов. Этот подход не решает проблему концептуальной эквивалентности окончательно, но делает её более явной и позволяет эмпирически оценить, насколько сильно связаны биологические, психологические и социальные компоненты стресса.
Теоретическое обоснование связи между индикаторами на разных уровнях является необходимым дополнением к статистическим методам и должно быть основано на понимании механизмов, связывающих уровни. Почему мы ожидаем связи между воспринимаемым стрессом и кортизолом? Теоретически, когнитивная оценка ситуации как угрожающей активирует миндалину и связанные с ней структуры, которые через гипоталамус запускают секрецию кортиколиберина и далее активацию гипоталамо-гипофизарно-надпочечниковой оси. Однако этот каскад модулируется множеством факторов: привыканием при повторяющихся стрессорах, индивидуальными различиями в реактивности оси, циркадными ритмами, сопутствующими условиями. Эти модулирующие факторы объясняют, почему связь между субъективным стрессом и кортизолом не является совершенной. Аналогичные механистические объяснения необходимы для понимания связи между средовыми стрессорами и субъективным переживанием, между биологическими изменениями и клиническими симптомами. Без такого теоретического понимания многоуровневое исследование остаётся эмпиристским сбором данных без объяснительной глубины.
Проблема операционализации имеет прямые следствия для шестого модуля курса, посвящённого измерению и биомаркерам стресса. Этот модуль систематически рассматривает методы измерения стресса на различных уровнях: субъективные методы, включающие шкалы воспринимаемого стресса, опросники жизненных событий, методы экологической моментальной оценки; физиологические методы, включающие измерение кортизола в различных биологических средах, оценку вариабельности сердечного ритма, кожно-гальванической реакции, нейровизуализацию; биомаркеры воспаления и метаболизма; маркеры биологического старения. Каждый из этих методов измеряет определённый аспект стресса со своими преимуществами и ограничениями, и грамотное исследование требует понимания того, что именно измеряет каждый метод и как интерпретировать согласование или расхождение между измерениями на разных уровнях. Освоение содержания шестого модуля предполагает понимание обсуждаемых здесь концептуальных проблем операционализации.
Практические рекомендации для многоуровневого исследования стресса, вытекающие из обсуждения проблемы операционализации, можно суммировать следующим образом. Во-первых, исследователь должен чётко определить, какой аспект стресса он изучает: объективные стрессоры среды, субъективное переживание, биологический ответ, долгосрочные последствия для здоровья, или связи между этими аспектами. Во-вторых, для каждого аспекта следует использовать множественные индикаторы, а не полагаться на единственное измерение. В-третьих, необходимо теоретически обосновать ожидаемые связи между индикаторами на разных уровнях и интерпретировать как согласование, так и расхождение в терминах механизмов. В-четвёртых, статистический анализ должен адекватно отражать структуру данных, используя методы латентно-переменного моделирования там, где это уместно. Наконец, интерпретация результатов должна учитывать ограничения каждого индикатора и избегать неоправданного обобщения от измеренного индикатора к целостному конструкту стресса.
Завершая обсуждение операционализации, следует подчеркнуть, что обсуждаемые трудности не являются специфическими для науки о стрессе, а отражают общую проблему измерения латентных конструктов в психологии, психиатрии и поведенческих науках. Интеллект, личностные черты, психопатологические синдромы, качество жизни — все эти конструкты сталкиваются с аналогичными проблемами операционализации и соотношения между конструктом и индикатором. Специфика стресса состоит в явно многоуровневой природе этого феномена, что делает проблему особенно острой. Однако накопленный в смежных областях опыт работы с латентными конструктами, развитые статистические методы и философская рефлексия над проблемами измерения предоставляют ресурсы для продуктивной работы с этими трудностями. Ключевым является сочетание концептуальной ясности, методологической строгости и теоретической глубины, которое позволяет превращать проблемы операционализации из препятствий в источники знания о сложной природе изучаемого феномена.
2.3 Статистические методы многоуровневого моделирования
Переход от концептуальных и методологических проблем к техническим аспектам анализа данных приводит нас к рассмотрению статистических методов, разработанных для работы с иерархически организованными данными, которые типичны для многоуровневых исследований стресса. Многоуровневое моделирование, также известное как иерархическое линейное моделирование или моделирование со смешанными эффектами, представляет собой семейство статистических методов, позволяющих одновременно анализировать вариацию на нескольких уровнях иерархии и оценивать как эффекты переменных на каждом уровне, так и взаимодействия между уровнями. Эти методы были разработаны в ответ на осознание того, что традиционные статистические подходы, предполагающие независимость наблюдений, неадекватны для данных, имеющих естественную иерархическую структуру, и их применение к таким данным ведёт к искажённым оценкам и ложным выводам. В исследованиях стресса иерархическая структура данных является скорее правилом, чем исключением, что делает владение многоуровневыми методами необходимой компетенцией для исследователя.
Типичная структура данных в исследовании организационного стресса иллюстрирует необходимость многоуровневых методов. Представим исследование, изучающее стресс и выгорание медицинских сестёр в сети больниц. Единицами анализа на первом уровне являются отдельные медсёстры, для каждой из которых измеряются индивидуальные характеристики: возраст, стаж, личностные черты, копинг-стратегии, воспринимаемый стресс, показатели выгорания. Однако медсёстры работают в отделениях, которые различаются по рабочей нагрузке, качеству руководства, атмосфере в коллективе, поддержке коллег. Отделения образуют второй уровень иерархии. Далее, отделения принадлежат больницам, которые различаются по организационной культуре, политикам управления персоналом, ресурсам. Больницы образуют третий уровень. Наконец, больницы могут быть сгруппированы по регионам с различной демографией, системой здравоохранения, экономическим положением, что создаёт четвёртый уровень. Стресс отдельной медсестры определяется факторами на всех этих уровнях, и адекватный анализ должен учитывать эту многоуровневую детерминацию.
Проблема зависимости наблюдений составляет техническое ядро обоснования многоуровневых методов и заслуживает детального рассмотрения. Традиционные статистические методы, такие как регрессионный анализ или дисперсионный анализ, предполагают, что наблюдения независимы друг от друга: информация об одном наблюдении не позволяет предсказать другое сверх того, что объясняется моделью. Однако в иерархических данных это предположение систематически нарушается. Медсёстры, работающие в одном отделении, подвержены влиянию одних и тех же условий труда, одного и того же руководителя, одной и той же атмосферы, и поэтому их показатели стресса более похожи друг на друга, чем показатели медсестёр из разных отделений. Это сходство внутри групп называется внутриклассовой корреляцией и может варьировать от нуля при полной независимости до единицы при полном совпадении показателей внутри группы. Игнорирование внутриклассовой корреляции при использовании традиционных методов приводит к занижению стандартных ошибок, завышению статистической значимости и, как следствие, к ложноположительным выводам о наличии эффектов.
Иерархическое линейное моделирование решает проблему зависимости наблюдений через явное разделение вариации на компоненты, относящиеся к разным уровням иерархии. В простейшем случае двухуровневой модели общая вариация в зависимой переменной разделяется на вариацию между группами и вариацию внутри групп. Вариация между группами отражает различия между отделениями или больницами в среднем уровне стресса сотрудников. Вариация внутри групп отражает различия между сотрудниками одного отделения. Предикторы могут вводиться на обоих уровнях: индивидуальные предикторы объясняют вариацию внутри групп, групповые предикторы объясняют вариацию между группами. Более того, многоуровневые модели позволяют оценивать, различаются ли эффекты индивидуальных предикторов между группами, то есть являются ли связи между переменными на индивидуальном уровне модерированными факторами группового уровня. Эта возможность моделирования кросс-уровневых взаимодействий делает многоуровневые методы мощным инструментом для изучения контекстуальных влияний на стресс.
Содержательный пример кросс-уровневого взаимодействия в исследовании стресса помогает понять практическую ценность многоуровневого моделирования. Предположим, мы изучаем связь между нагрузкой и выгоранием у медсестёр. На индивидуальном уровне можно ожидать, что медсёстры с более высокой субъективно воспринимаемой нагрузкой будут иметь более высокое выгорание. Однако эта связь может зависеть от характеристик отделения: в отделениях с высокой социальной поддержкой связь между нагрузкой и выгоранием может быть слабее, поскольку поддержка коллег буферирует эффект нагрузки. Это и есть кросс-уровневое взаимодействие: переменная группового уровня модерирует связь между переменными индивидуального уровня. Традиционный регрессионный анализ не способен адекватно оценить такое взаимодействие; более того, попытка включить групповые переменные в индивидуальную регрессию приводит к статистическим артефактам. Многоуровневое моделирование позволяет корректно оценить как основные эффекты на обоих уровнях, так и их взаимодействие, предоставляя инструмент для изучения того, как контекст модифицирует индивидуальные связи.
Технические требования к применению многоуровневого моделирования создают практические ограничения, которые исследователь должен учитывать при планировании исследования. Первое и наиболее существенное требование касается размера выборки на каждом уровне иерархии. Для надёжной оценки эффектов на групповом уровне необходимо достаточное количество групп; общая рекомендация составляет не менее тридцати, а предпочтительно пятидесяти групп для оценки межгрупповых эффектов и кросс-уровневых взаимодействий. Количество наблюдений внутри групп также должно быть достаточным, хотя здесь требования менее строги и зависят от величины внутриклассовой корреляции. Эти требования означают, что полноценное многоуровневое исследование организационного стресса требует данных из десятков организаций с сотнями или тысячами участников, что создаёт значительные логистические и финансовые барьеры. Второе требование касается корректной спецификации модели: неправильное определение уровней, ошибочное приписывание переменных к уровням, пропуск значимых предикторов могут привести к искажённым выводам.
Многоуровневые структурные уравнения расширяют возможности иерархического линейного моделирования, интегрируя его с методами моделирования латентных переменных. В традиционном иерархическом линейном моделировании все переменные являются наблюдаемыми, то есть непосредственно измеренными. Многоуровневое моделирование структурными уравнениями позволяет моделировать латентные переменные на каждом уровне иерархии, что особенно ценно с учётом обсуждавшихся ранее проблем операционализации. Если воспринимаемый стресс измеряется несколькими индикаторами, можно построить латентный фактор воспринимаемого стресса и анализировать связи этого фактора с предикторами и исходами, отделяя истинную вариацию в конструкте от ошибки измерения. Более того, можно анализировать структуру латентных факторов на разных уровнях: латентный фактор на индивидуальном уровне может отражать индивидуальные различия в стрессе, а латентный фактор на групповом уровне — межгрупповые различия в климате или культуре. Эта методологическая изощрённость требует ещё более строгих требований к размеру выборки и компетенции исследователя.
Альтернативные и дополнительные подходы к анализу многоуровневых данных включают байесовские методы и методы машинного обучения, каждый со своими преимуществами. Байесовские многоуровневые модели предлагают ряд преимуществ по сравнению с частотными подходами, включая возможность работы с меньшими выборками через информативные априорные распределения, более гибкое моделирование сложных структур данных, естественную оценку неопределённости параметров. В контексте стресса байесовские методы особенно полезны, когда данные ограничены, но имеется существенная теоретическая информация из предшествующих исследований. Методы машинного обучения, такие как случайные леса или градиентный бустинг с учётом иерархической структуры, позволяют выявлять сложные нелинейные связи и взаимодействия, которые могут быть упущены линейными моделями. Однако методы машинного обучения ориентированы на предсказание, а не на объяснение, и их результаты труднее интерпретировать в терминах каузальных механизмов. Выбор между подходами определяется исследовательской целью: объяснение механизмов, предсказание исходов или построение классификаций.
Связь обсуждаемых статистических методов с содержанием курса проявляется в нескольких точках. Шестой модуль, посвящённый измерению, рассматривает не только инструменты сбора данных, но и базовые принципы их анализа, включая понимание иерархической структуры и необходимости адекватных статистических методов. Десятый модуль о контекстах и популяциях непосредственно оперирует многоуровневыми конструкциями: профессиональный контекст, семейный контекст, культурный контекст представляют собой уровни, на которых формируются условия для индивидуального стресса. Модуль о профессиональном выгорании обсуждает организационные факторы, которые методологически корректно анализируются только многоуровневыми методами. Одиннадцатый модуль о передовых исследованиях рассматривает современные методологические разработки, включая продвинутые многоуровневые модели и их интеграцию с вычислительными подходами. Понимание логики и возможностей многоуровневого моделирования является необходимым для критической оценки литературы и для планирования собственных исследований.
Ограничения многоуровневого моделирования следует понимать не как повод для отказа от этих методов, а как условия их грамотного применения. Первое ограничение касается предположения о правильной спецификации модели: если важные предикторы пропущены или функциональная форма связей неправильна, оценки будут смещены независимо от статистической изощрённости метода. Второе ограничение связано с корреляционной природой большинства многоуровневых исследований: даже корректно оценённые связи между переменными на разных уровнях не доказывают причинность. Третье ограничение касается генерализуемости: выводы о кросс-уровневых взаимодействиях, полученные в одной популяции или культуре, могут не воспроизводиться в других контекстах. Эти ограничения не специфичны для многоуровневых методов, а отражают общие проблемы наблюдательных исследований. Многоуровневое моделирование предоставляет инструменты для корректной оценки связей в иерархических данных, но интерпретация этих связей требует теоретического понимания и дополнительных источников доказательств.
2.4 Проблема временной динамики: лонгитюдный и кросс-секционный подходы
Стресс по своей сущности является процессом, разворачивающимся во времени, и адекватное его изучение требует методов, способных схватывать эту темпоральную природу, однако значительная часть эмпирических исследований стресса основана на кросс-секционных дизайнах, которые предоставляют лишь моментальный срез и не позволяют наблюдать динамику изменений. Кросс-секционное исследование собирает данные от группы участников в один момент времени и анализирует связи между переменными, измеренными одновременно. Хотя такие исследования могут выявить корреляции и ассоциации, они принципиально неспособны установить временной порядок событий: если обнаружена связь между воспринимаемым стрессом и депрессивными симптомами, невозможно определить, предшествует ли стресс депрессии, депрессия стрессу, или обе переменные являются следствиями некоторой третьей причины. Эта неспособность к установлению временного порядка резко ограничивает возможности каузальных выводов, которые критичны для понимания механизмов развития стресс-связанных расстройств и для разработки интервенций.
Лонгитюдные исследования, отслеживающие одних и тех же участников на протяжении времени, предоставляют существенные преимущества для изучения динамики стресса. Простейший лонгитюдный дизайн включает два измерения: исходное и последующее, разделённые интервалом времени. Если исходный уровень воспринимаемого стресса предсказывает последующее развитие депрессивных симптомов при контроле исходного уровня депрессии, это даёт более сильное основание для вывода о том, что стресс способствует развитию депрессии, нежели простая кросс-секционная корреляция. Более сложные лонгитюдные дизайны включают множественные волны измерений, позволяющие моделировать траектории изменения переменных во времени и анализировать, как изменения в одной переменной связаны с последующими изменениями в другой. Панельные модели с перекрёстными лагами позволяют одновременно оценивать влияние стресса на депрессию и депрессии на стресс в последовательных волнах измерений, выявляя направленность связей. Моделирование латентных кривых роста позволяет описывать индивидуальные траектории изменения и анализировать предикторы различий в этих траекториях.
Применение лонгитюдных дизайнов к многоуровневому исследованию стресса создаёт сложные структуры данных, требующие специализированных аналитических подходов. Если мы отслеживаем сотрудников организаций на протяжении нескольких лет, данные имеют структуру «наблюдения в рамках индивидов в рамках организаций в рамках временных точек». Время само становится уровнем иерархии, и повторные измерения от одного индивида не независимы, что требует многоуровневых методов с учётом вложенности временных точек в индивидов. Более того, индивиды, организации и их характеристики могут меняться со временем: сотрудник может сменить должность или отделение, организация может изменить политику, внешний контекст может трансформироваться. Моделирование этих изменений требует динамических многоуровневых моделей, которые позволяют переменным на всех уровнях изменяться во времени. Такие модели технически сложны, требуют больших выборок с длительным наблюдением и высокой компетенции исследователя, но только они способны адекватно отразить динамическую многоуровневую природу стресса.
Метод экологической моментальной оценки представляет собой особый подход к интенсивному лонгитюдному изучению стресса в естественных условиях жизни участников. В этом методе участники многократно в течение дня отвечают на краткие вопросы о своём состоянии, деятельности, социальном контексте, используя мобильные устройства. Типичный протокол может включать пять-десять моментальных оценок в день на протяжении одной-двух недель, генерируя сотни наблюдений от каждого участника. Эти данные позволяют изучать динамику стресса с высоким временным разрешением: как настроение и стресс флуктуируют в течение дня, как они связаны с событиями и контекстами, как паттерны дневной динамики предсказывают долгосрочные исходы. Добавление физиологических измерений через носимые устройства — мониторы сердечного ритма, датчики активности, даже непрерывное измерение кортизола — позволяет синхронно отслеживать субъективные переживания и биологические маркеры, изучая их динамическую связь в реальном времени. Этот подход особенно ценен для изучения эмерджентных паттернов: как последовательности микрострессоров накапливаются и приводят к состояниям истощения, как восстановительные процессы разворачиваются после стрессовых эпизодов.
Каузальные преимущества лонгитюдных дизайнов основаны на принципе темпорального приоритета: причина должна предшествовать следствию во времени. Если изменение в переменной A в момент времени T1 предшествует и предсказывает изменение в переменной B в момент T2, это даёт основания рассматривать A как потенциальную причину B. Однако лонгитюдные данные сами по себе не гарантируют каузальных выводов, поскольку остаётся возможность конфаундинга: некоторая третья переменная может вызывать изменения и в A, и в B с различным временным лагом, создавая видимость причинной связи между A и B. Контроль измеренных конфаундеров в лонгитюдном анализе повышает уверенность в каузальных интерпретациях, но не устраняет проблему полностью, поскольку всегда могут существовать неизмеренные конфаундеры. Более того, выбор временных интервалов между измерениями влияет на выводы: если интервал слишком короток, эффект может не успеть проявиться; если слишком длинен, он может быть опосредован или затушёван другими процессами. Оптимальный интервал должен соответствовать теоретически ожидаемому времени развития эффекта, которое часто неизвестно.
Интеграция лонгитюдных дизайнов с многоуровневыми биологическими измерениями открывает возможности для изучения того, как динамика психологического стресса связана с динамикой биологических изменений. Представим исследование, в котором участники ежедневно сообщают о своём стрессе и настроении, еженедельно предоставляют образцы слюны для измерения кортизола и воспалительных маркеров, и ежемесячно проходят расширенное обследование с оценкой функционального состояния. Такой дизайн позволяет задать вопросы: предсказывает ли накопление дневного стресса за неделю повышение воспалительных маркеров в конце недели? Предсказывает ли устойчивое повышение воспаления ухудшение настроения и когнитивных функций в последующие недели? Как индивидуальные траектории стресса связаны с траекториями биологических изменений? Эти вопросы требуют данных с множественными временными разрешениями и с измерениями на разных уровнях, что создаёт значительные логистические сложности, но предоставляет уникальные возможности для понимания динамических связей между уровнями.
Проблемы лонгитюдных исследований включают не только технические сложности, но и практические ограничения, которые объясняют сохраняющееся преобладание кросс-секционных дизайнов в литературе. Атритция, то есть выбывание участников из исследования с течением времени, создаёт систематическое смещение, если выбывание связано с изучаемыми переменными: наиболее стрессированные участники могут с большей вероятностью прекращать участие, что занижает оценки стресса в оставшейся выборке. Реактивность измерений означает, что сам процесс повторного измерения может изменять изучаемое явление: участники могут становиться более осведомлёнными о своём стрессе, изменять поведение или уставать от исследования. Стоимость и продолжительность лонгитюдных исследований существенно превышают кросс-секционные, что ограничивает их применение при ограниченном финансировании. Наконец, быстрое устаревание инструментов измерения при длительных исследованиях создаёт проблемы сопоставимости данных, собранных в разные периоды. Эти ограничения не обесценивают лонгитюдный подход, но требуют тщательного планирования для их минимизации.
Методологические стратегии работы с проблемами лонгитюдных исследований включают планирование избыточных выборок с учётом ожидаемой атритции, использование статистических методов для оценки и коррекции смещения, связанного с выбыванием, тщательный выбор временных интервалов на основе теоретических соображений о темпоральности изучаемых процессов. Для интенсивных лонгитюдных исследований с моментальной оценкой важно балансировать информативность частых измерений и бремя для участников, которое ведёт к усталости и снижению качества ответов. Современные технологии пассивного сбора данных через смартфоны и носимые устройства — геолокация, паттерны использования телефона, биометрика — позволяют собирать данные без активного участия респондентов, снижая бремя и потенциальную реактивность. Однако эти методы поднимают этические вопросы приватности и информированного согласия, которые требуют внимательного рассмотрения.
Связь проблемы временной динамики с содержанием курса проявляется в нескольких аспектах. Концепция аллостатической нагрузки, центральная для первого модуля, принципиально темпоральна: она описывает накопление физиологического износа с течением времени в ответ на хронические стрессоры. Понимание развития выгорания в восьмом модуле требует лонгитюдной перспективы: выгорание не возникает мгновенно, а формируется через фазы, и понимание этих фаз требует отслеживания траекторий. Эффективность интервенций, рассматриваемая в девятом модуле, определяется их способностью изменять траектории стресса и его последствий во времени, что требует лонгитюдной оценки исходов. Шестой модуль о методах измерения включает обсуждение экологической моментальной оценки как метода интенсивного лонгитюдного измерения. Одиннадцатый модуль о передовых исследованиях рассматривает динамические системные модели, которые математически формализуют темпоральную динамику стресса и позволяют моделировать критические переходы и точки бифуркации.
Завершая обсуждение временной динамики, важно подчеркнуть, что кросс-секционные и лонгитюдные дизайны не взаимоисключающи, а дополняют друг друга в программе исследований. Кросс-секционные исследования эффективны для первичного выявления связей между переменными, для изучения индивидуальных различий в определённый момент, для исследований с большими выборками и редкими явлениями. Они также служат основой для планирования лонгитюдных исследований, определяя релевантные переменные и гипотезы. Лонгитюдные исследования необходимы для проверки каузальных гипотез, для изучения траекторий и механизмов изменения, для понимания динамических связей между переменными на разных уровнях. Оптимальная исследовательская программа включает оба типа дизайнов, используя сильные стороны каждого для последовательного накопления знания о многоуровневой динамике стресса.
2.5 Интеграция разнородных данных: омиксные подходы
Революция в технологиях молекулярной биологии последних десятилетий создала возможности для одновременного измерения тысяч и миллионов переменных на молекулярном уровне, что породило семейство омиксных подходов и поставило перед наукой о стрессе беспрецедентные вызовы и возможности интеграции разнородных данных. Геномика изучает последовательность и вариации ДНК, включая полногеномные ассоциативные исследования, связывающие генетические варианты с фенотипами стресс-связанных расстройств. Эпигеномика исследует химические модификации ДНК и гистонов, которые регулируют экспрессию генов без изменения последовательности и могут отражать влияние стрессового опыта на молекулярном уровне. Транскриптомика измеряет экспрессию тысяч генов через количественную оценку соответствующих молекул РНК, создавая транскрипционный профиль клетки или ткани в ответ на стресс. Протеомика идентифицирует и количественно оценивает белки, являющиеся конечными продуктами экспрессии генов и непосредственными исполнителями клеточных функций. Метаболомика измеряет малые молекулы — метаболиты, являющиеся субстратами и продуктами биохимических реакций и отражающие текущее метаболическое состояние организма. Микробиомика характеризует состав и функции микробных сообществ, прежде всего кишечной микробиоты, которая, как становится всё более ясно, играет существенную роль в регуляции стресса и настроения через ось кишечник-мозг.
Каждый из омиксных подходов генерирует массивы данных беспрецедентного размера и сложности, что создаёт специфические методологические вызовы анализа. Типичное транскриптомное исследование измеряет экспрессию двадцати-тридцати тысяч генов у каждого участника, создавая многомерное пространство данных, в котором количество переменных многократно превышает количество наблюдений. Эта ситуация, известная как «проклятие размерности», делает неприменимыми традиционные статистические методы и порождает проблему множественных сравнений: при тестировании тысяч гипотез сотни могут оказаться «значимыми» просто по случайности. Методы коррекции множественных сравнений, такие как поправка Бонферрони или контроль доли ложных открытий, снижают ложноположительные результаты, но одновременно повышают риск пропуска реальных эффектов. Альтернативные подходы включают использование априорных знаний для группировки генов в биологически осмысленные наборы и тестирования обогащённости этих наборов вместо отдельных генов, а также методы машинного обучения для выявления паттернов в высокоразмерных данных. Однако результаты машинного обучения требуют внешней валидации и часто труднообъяснимы биологически.
Мультиомиксная интеграция представляет собой попытку связать данные с разных омиксных платформ для получения более полной картины молекулярных механизмов стресса. Логика интеграции основана на понимании того, что информация течёт от генома через транскриптом и протеом к метаболому и в конечном счёте к фенотипу: генетические варианты влияют на экспрессию генов, экспрессия определяет уровни белков, белки катализируют метаболические реакции, метаболиты влияют на клеточные функции. Изучая связи между омиксными уровнями, можно реконструировать молекулярные пути, через которые генетическая вариация транслируется в фенотипическое разнообразие ответов на стресс. Например, генетический вариант в гене FKBP5, ассоциированный с повышенным риском посттравматического расстройства, влияет на экспрессию этого гена, продукт которого регулирует чувствительность глюкокортикоидного рецептора, что изменяет паттерн транскрипционного ответа на кортизол и, в конечном счёте, метаболические и функциональные последствия стресса. Прослеживание такой цепочки требует интеграции геномных, транскриптомных и функциональных данных.
Методы мультиомиксной интеграции включают несколько подходов с различными допущениями и целями. Сетевой анализ строит графы, в которых узлы представляют молекулы, а рёбра — их взаимодействия или корреляции, и ищет модули взаимосвязанных молекул, которые могут представлять функциональные единицы. Связывание модулей из разных омиксных слоёв позволяет идентифицировать молекулярные подписи стресса, охватывающие несколько уровней. Методы снижения размерности, такие как канонический корреляционный анализ или его современные расширения, ищут латентные факторы, объясняющие ковариацию между омиксными платформами, выделяя общие источники вариации. Методы машинного обучения, включая тензорную факторизацию и глубокие нейронные сети, способны обнаруживать сложные нелинейные паттерны в интегрированных данных, хотя интерпретируемость их результатов остаётся проблемой. Байесовские иерархические модели позволяют формализовать априорные знания о структуре связей между омиксными уровнями и оценивать неопределённость выводов, что особенно ценно при ограниченных размерах выборок.
Конкретный пример мультиомиксного подхода к стрессу иллюстрирует возможности и сложности интеграции. Предположим, исследователь стремится выявить биотипы депрессии с различными молекулярными механизмами. Он собирает образцы крови от большой когорты пациентов с депрессией и здоровых контролей, проводит генотипирование для оценки генетического риска, транскриптомное профилирование для оценки экспрессии генов, метаболомное профилирование для оценки метаболического состояния, параллельно собирая детальные клинические и психологические данные. Интегративный анализ может выявить кластер пациентов с высоким полигенным риском воспалительных заболеваний, повышенной экспрессией провоспалительных генов, метаболическим профилем, характерным для системного воспаления, и клинической картиной с преобладанием соматических симптомов депрессии, таких как усталость, нарушения сна и аппетита. Другой кластер может характеризоваться генетическим риском в области нейропластичности, сниженной экспрессией генов, связанных с синаптической функцией, метаболическими маркерами нарушенного энергетического обмена и клинической картиной с преобладанием когнитивных симптомов. Третий кластер может демонстрировать дисрегуляцию генов циркадных ритмов, соответствующие метаболические нарушения и выраженные нарушения сна и суточной ритмики. Каждый из этих биотипов представляет собой различную реализацию клинической депрессии на молекулярном уровне и потенциально требует различных терапевтических подходов. Такой анализ воплощает идею множественной реализуемости, обсуждавшуюся в философском разделе, в конкретную эмпирическую программу и открывает путь к персонализированной медицине стресс-связанных расстройств.
Вычислительные и статистические сложности мультиомиксной интеграции требуют специализированной инфраструктуры и экспертизы, которые выходят за рамки традиционной подготовки исследователей стресса. Объёмы данных измеряются гигабайтами и терабайтами, требуя высокопроизводительных вычислительных кластеров для хранения и обработки. Анализ включает десятки программных инструментов и конвейеров обработки, каждый со своими параметрами и допущениями, и результаты могут существенно зависеть от выбора методов и настроек. Воспроизводимость анализа требует тщательного документирования всех шагов обработки, версий программного обеспечения, параметров алгоритмов. Проблема множественных сравнений усугубляется при интеграции нескольких омиксных платформ, где тестируются не только тысячи отдельных молекул, но и миллионы потенциальных связей между молекулами разных типов. Валидация результатов требует независимых когорт, экспериментальных подтверждений функциональной значимости выявленных паттернов, биологической интерпретации статистически выявленных ассоциаций. Всё это создаёт барьеры для входа и требует междисциплинарных команд, объединяющих молекулярных биологов, биоинформатиков, статистиков, клиницистов и психологов.
Интерпретируемость результатов мультиомиксного анализа остаётся одной из центральных проблем, особенно при использовании методов машинного обучения. Алгоритм может выявить паттерн в данных, предсказывающий клинический исход с высокой точностью, но этот паттерн может включать сотни переменных со сложными нелинейными взаимодействиями, которые невозможно свести к простому биологическому объяснению. Такие модели, иногда называемые «чёрными ящиками», имеют предсказательную ценность, но ограниченную объяснительную силу. Для науки о стрессе, стремящейся понять механизмы, а не только предсказывать исходы, это создаёт напряжение между предсказательной мощью и биологической интерпретируемостью. Современные разработки в области объяснимого искусственного интеллекта предлагают методы извлечения интерпретируемых правил из сложных моделей, но эти методы несовершенны и не устраняют проблему полностью. Прагматическое решение состоит в использовании машинного обучения для генерации гипотез, которые затем валидируются экспериментально и интерпретируются биологически через традиционные механистические исследования.
Связь мультиомиксных подходов с содержанием курса проявляется прежде всего в материале о биологических уровнях анализа стресса. Первый модуль о клеточных и молекулярных механизмах закладывает концептуальную основу для понимания того, что измеряют омиксные методы: экспрессия генов, белковые продукты, метаболические пути. Четвёртый модуль об иммунитете, воспалении и метаболизме непосредственно связан с омиксными исследованиями воспалительных и метаболических сигнатур стресса. Шестой модуль о биомаркерах включает обсуждение мультиомиксных подходов как передовых методов характеризации биологического стресс-ответа. Одиннадцатый модуль о передовых исследованиях рассматривает интеграцию омиксных данных как одно из ключевых направлений развития науки о стрессе. Понимание возможностей и ограничений этих подходов необходимо для критической оценки современной литературы и для осознанного участия в междисциплинарных исследовательских проектах.
Перспективы развития мультиомиксных исследований стресса связаны с преодолением текущих ограничений через технологические инновации, методологические разработки и накопление данных. Технологии одноклеточного секвенирования позволяют изучать молекулярные профили отдельных клеток, а не усреднённые сигналы от смеси клеточных типов, что существенно повышает разрешающую способность и позволяет идентифицировать специфические клеточные популяции, ответственные за стрессовый ответ. Пространственная транскриптомика добавляет информацию о локализации экспрессии в тканях, связывая молекулярные данные с анатомической структурой. Лонгитюдные омиксные исследования позволяют отслеживать молекулярные траектории, а не только срезы, интегрируя временную динамику, обсуждавшуюся в предыдущем разделе. Международные консорциумы и базы данных накапливают омиксные данные от сотен тысяч участников, создавая ресурсы для мета-анализа и валидации. Все эти развития обещают более глубокое понимание молекулярных механизмов стресса, хотя полная реализация этого обещания остаётся делом будущего.
Завершая обсуждение мультиомиксной интеграции, необходимо подчеркнуть, что эти подходы не заменяют, а дополняют традиционные методы исследования стресса. Омиксные данные предоставляют беспрецедентную детализацию молекулярного уровня, но они не охватывают системный, психологический и социальный уровни, которые столь же существенны для понимания стресса. Молекулярная подпись стресса в крови ничего не говорит о субъективном переживании человека, о смыслах, которые он приписывает своему опыту, о социальных условиях, породивших стрессор. Интегративное понимание стресса требует связывания омиксных данных с измерениями на других уровнях, что возвращает нас к проблемам многоуровневого моделирования, обсуждавшимся в предыдущих разделах. Мультиомиксная интеграция представляет собой важный, но не единственный компонент этого интегративного предприятия.
2.6 Причинность в многоуровневых системах: методы каузальной инференции
Установление причинно-следственных связей представляет собой центральную цель научного познания, и в многоуровневых исследованиях стресса эта задача приобретает особую сложность, обусловленную переплетением факторов на различных уровнях организации, многонаправленностью влияний и ограниченными возможностями экспериментального контроля. Понимание причин стресса и его последствий необходимо не только для теоретического осмысления феномена, но и для практических целей профилактики и терапии: чтобы эффективно вмешиваться, нужно знать, какие факторы являются причинами, а какие лишь коррелятами или следствиями. Корреляционные исследования, составляющие большую часть эмпирической литературы по стрессу, могут выявить связи между переменными, но не могут определить их каузальный статус. Хронический стресс коррелирует с депрессией, но является ли он причиной депрессии, следствием продромальных симптомов, или обе переменные являются следствиями общей причины? Низкий социально-экономический статус ассоциирован с повышенной аллостатической нагрузкой, но является ли бедность причиной биологического износа, или оба фактора отражают третью переменную, такую как генетическая предрасположенность или характеристики среды раннего развития? Эти вопросы имеют не только теоретическое значение, но и прямые практические следствия для политики и интервенций.
Рандомизированное контролируемое испытание остаётся золотым стандартом для установления причинности благодаря своей способности устранять проблему конфаундинга через случайное распределение участников по условиям. Когда участники случайным образом назначаются в экспериментальную или контрольную группу, любые систематические различия между группами, кроме экспериментального воздействия, элиминируются, и наблюдаемые различия в исходах могут быть атрибутированы воздействию. Однако возможности экспериментального подхода в исследованиях стресса существенно ограничены этическими, практическими и концептуальными соображениями. Этически невозможно случайным образом назначать людей на воздействие хроническим стрессом, травмой, социальным исключением или бедностью для изучения их последствий. Практически невозможно экспериментально манипулировать многими интересующими переменными, такими как генетические варианты, ранний опыт или социально-экономический статус. Концептуально сложно создать в лаборатории стрессоры, эквивалентные по своему воздействию реальным жизненным ситуациям; лабораторные стрессоры, такие как публичное выступление или арифметические задачи под давлением, вызывают острый кратковременный стресс, который качественно отличается от хронического стресса реальной жизни. Эти ограничения означают, что для многих ключевых вопросов науки о стрессе экспериментальные данные принципиально недоступны, и исследователи вынуждены полагаться на наблюдательные данные с их ограничениями для каузальных выводов.
Менделевская рандомизация представляет собой остроумный методологический подход, который использует генетические варианты как инструментальные переменные для каузальной инференции в наблюдательных данных. Логика метода основана на нескольких ключевых предположениях. Во-первых, генетические варианты определяются при зачатии и не могут быть следствием изучаемых исходов, что устанавливает временной приоритет. Во-вторых, распределение генетических вариантов в популяции приблизительно случайно относительно средовых конфаундеров, что создаёт естественный аналог рандомизации. В-третьих, если генетический вариант влияет на исход только через экспозицию, он может служить инструментом для оценки каузального эффекта экспозиции на исход. Рассмотрим пример: для изучения каузального эффекта образования на стресс и здоровье можно использовать генетические варианты, ассоциированные с продолжительностью обучения. Если люди с генетической предрасположенностью к более длительному образованию имеют лучшие показатели здоровья, и этот эффект опосредован фактическим уровнем образования, это даёт основания для каузальной интерпретации связи между образованием и здоровьем. Менделевская рандомизация широко применяется в эпидемиологии и начинает использоваться в исследованиях стресса для проверки каузального статуса различных факторов риска.
Ограничения менделевской рандомизации требуют осторожности в интерпретации её результатов и понимания условий, при которых метод даёт надёжные выводы. Первое критическое предположение состоит в отсутствии плейотропии: генетический вариант должен влиять на исход только через изучаемую экспозицию, а не через другие пути. Если вариант, ассоциированный с образованием, также влияет на когнитивные способности, которые независимо влияют на здоровье, каузальная оценка будет искажена. Современные методы, такие как взвешивание по медиане или отношение правдоподобия, пытаются справиться с плейотропией, но не устраняют проблему полностью. Второе предположение касается силы инструмента: генетические варианты должны достаточно сильно предсказывать экспозицию, иначе оценки будут неточны и смещены. Для многих интересующих экспозиций генетические предикторы слабы, что ограничивает применимость метода. Третье ограничение связано с популяционной стратификацией: если генетические варианты и исходы различаются между субпопуляциями по причинам, не связанным с каузальным эффектом, это создаёт ложные ассоциации. Эти ограничения не обесценивают метод, но требуют тщательной проверки предположений и триангуляции с другими источниками доказательств.
Естественные эксперименты представляют собой ещё один подход к каузальной инференции в наблюдательных данных, использующий ситуации, в которых внешние события создают квазиэкспериментальные условия. Когда изменение социальной политики, экономическое потрясение, природная катастрофа или другое событие случайно или квазислучайно воздействует на часть популяции, это создаёт возможность сравнить исходы у затронутых и не затронутых групп, приближаясь к логике экспериментального дизайна. Классический пример в исследованиях стресса: изучение последствий террористических атак одиннадцатого сентября для психического здоровья позволяет сравнивать население Нью-Йорка, непосредственно затронутое событиями, с населением отдалённых регионов, предполагая, что до события группы были сопоставимы. Изменения в политике социального обеспечения, минимальной заработной плате, доступе к здравоохранению создают естественные эксперименты для изучения каузальных эффектов социально-экономических факторов на стресс и здоровье. Метод разрывной регрессии использует пороги допуска к программам или льготам: люди чуть выше и чуть ниже порога сопоставимы, но различаются по экспозиции, что позволяет оценить каузальный эффект. Метод разности разностей сравнивает изменения в исходах до и после интервенции между группами, получившими и не получившими воздействие.
Каузальное моделирование с использованием направленных ациклических графов предоставляет формальный язык для представления каузальных предположений и определения условий, при которых наблюдательные данные позволяют идентифицировать каузальные эффекты. Направленный ациклический граф представляет собой диаграмму, в которой узлы обозначают переменные, а направленные рёбра — каузальные связи между ними. Граф эксплицирует предположения исследователя о каузальной структуре изучаемой системы: какие переменные влияют на какие, какие связи прямые, какие опосредованные. На основе графа можно формально определить, какие переменные необходимо контролировать для идентификации каузального эффекта одной переменной на другую, а какие контролировать нельзя, поскольку это приведёт к смещению. Критерии бэкдор и фронтдор формализуют условия, при которых каузальный эффект идентифицируем. Этот подход не даёт новых данных, но дисциплинирует мышление о каузальности и делает предположения явными, что позволяет их критически оценивать. В исследованиях стресса каузальные графы помогают структурировать сложные многоуровневые связи и определять, какие измерения необходимы для тестирования каузальных гипотез.
Применение каузального моделирования к многоуровневым системам стресса наталкивается на специфические сложности, связанные с множественностью уровней и направлений причинности. Каузальные связи в системе стресса не являются однонаправленными: биологические факторы влияют на психологические состояния, но психологические состояния через нисходящую причинность влияют на биологию; индивидуальные характеристики формируются социальным контекстом, но индивиды также формируют свой социальный контекст через выбор и поведение. Эти взаимные влияния и петли обратной связи создают сложные каузальные структуры, которые трудно представить ациклическими графами и для которых стандартные методы идентификации не всегда применимы. Динамические каузальные модели, учитывающие временную структуру взаимных влияний, и агентные модели, симулирующие взаимодействия на различных уровнях, предлагают альтернативные подходы к моделированию таких систем. Однако эти методы требуют более сильных допущений и больших объёмов данных, а их результаты чувствительны к спецификации модели. Каузальная инференция в сложных многоуровневых системах остаётся на переднем крае методологических разработок.
Триангуляция доказательств представляет собой стратегию повышения уверенности в каузальных выводах через интеграцию данных из разных источников, полученных разными методами и основанных на разных допущениях. Если менделевская рандомизация, естественный эксперимент и лонгитюдное наблюдательное исследование с контролем конфаундеров приводят к согласующимся выводам о каузальном эффекте, уверенность в каузальности существенно повышается, поскольку маловероятно, что все три метода систематически искажены в одном направлении. Различные методы имеют различные паттерны возможного смещения: менделевская рандомизация уязвима к плейотропии, естественные эксперименты — к нарушению предположения о случайности распределения, наблюдательные исследования — к неизмеренному конфаундингу. Когда выводы сходятся несмотря на эти различные уязвимости, вероятность того, что все методы ошибочны, снижается. Принцип триангуляции особенно важен в науке о стрессе, где экспериментальные данные ограничены и каузальные выводы неизбежно основываются на комбинации косвенных источников доказательств.
Практические следствия обсуждения каузальности для исследования и практики многообразны. Для исследователя осознание различия между корреляцией и причинностью определяет выбор дизайна, методов и осторожность в формулировках. Утверждение «хронический стресс вызывает депрессию» требует каузального обоснования, которое не может быть предоставлено одним кросс-секционным корреляционным исследованием, каким бы большим и детальным оно ни было. Для клинициста понимание каузального статуса связей определяет ожидания от интервенций: воздействие на истинную причину должно изменить следствие, воздействие на коррелят или следствие не обязательно изменит причину. Для политика каузальные знания определяют, какие вмешательства на популяционном уровне будут эффективны: если социально-экономическое неравенство является причиной хронического стресса и связанных с ним заболеваний, то политики редукции неравенства должны улучшить здоровье населения; если связь лишь корреляционная, такие политики могут не достичь цели. Эти практические ставки делают методологию каузальной инференции не абстрактным упражнением, а критическим инструментом для основанных на доказательствах решений.
Связь проблематики причинности с содержанием курса проявляется на протяжении всех модулей. Первый модуль о механизмах стресса по существу представляет каузальные модели того, как стрессоры вызывают биологические изменения. Модули о патологии описывают каузальные пути, ведущие от хронического стресса к заболеваниям различных систем. Модуль об интервенциях оценивает каузальную эффективность различных методов помощи, что требует рандомизированных испытаний или их аналогов. Нулевой модуль о научной грамотности закладывает понимание различия между корреляцией и причинностью как базовый элемент критического мышления. Одиннадцатый модуль о передовых исследованиях рассматривает новые подходы к каузальной инференции, включая вычислительное каузальное моделирование и системную биологию. Сквозная тема каузальности объединяет разрозненные факты в целостное понимание механизмов стресса и его последствий.
Завершая обсуждение причинности в многоуровневых системах, необходимо признать принципиальную неполноту любых каузальных знаний о таких сложных феноменах, как стресс, и важность эпистемической скромности в формулировании выводов. Даже наилучшие методы каузальной инференции предоставляют лишь вероятностные оценки, основанные на предположениях, которые никогда не могут быть полностью верифицированы. Сложность многоуровневых систем означает, что всегда остаются неучтённые факторы, неизмеренные конфаундеры, неспецифицированные взаимодействия. Это не повод для агностицизма или отказа от каузальных исследований; это призыв к осторожности, триангуляции, накоплению доказательств из различных источников и готовности пересматривать выводы при появлении новых данных. Наука о стрессе, как и любая наука о сложных системах, продвигается через последовательное приближение к пониманию, а не через окончательные и безоговорочные истины, и методологическая рефлексия над условиями и ограничениями каузального знания составляет неотъемлемую часть этого продвижения.
3. Трансляционный разрыв: от фундаментальных исследований к клинической практике
3.1 Феномен трансляционного разрыва и его масштаб
Переход от фундаментальных научных открытий к практическим клиническим приложениям представляет собой одну из наиболее критических и одновременно проблематичных фаз в развитии биомедицинского знания, и в области стресс-связанных расстройств этот переход сопряжён с особенно выраженными трудностями, которые получили название трансляционного разрыва. Этот феномен характеризует систематическое расхождение между многообещающими результатами доклинических исследований и их неспособностью воспроизводиться в клинических условиях, между потенциалом, демонстрируемым на уровне клеточных культур и животных моделей, и разочаровывающими исходами терапевтических испытаний на людях. Метафора «долины смерти», используемая для описания этого разрыва, отражает тот факт, что подавляющее большинство потенциальных терапевтических агентов, успешно преодолевших доклиническую фазу, погибает при попытке пересечь пространство между лабораторией и клиникой. Признание масштаба и глубины трансляционного разрыва не означает критики научного метода или обесценивания фундаментальных исследований; напротив, оно представляет собой необходимую рефлексию над методологическими ограничениями и условие для разработки стратегий их преодоления.
Статистические данные о масштабе трансляционного разрыва в биомедицине в целом и в психофармакологии в частности рисуют картину систематической неэффективности трансляционного процесса. По различным оценкам, от восьмидесяти до девяноста процентов потенциальных лекарственных препаратов, демонстрирующих эффективность на доклинической стадии, не подтверждают своей эффективности в клинических испытаниях на людях. Это означает, что лишь один из десяти или даже меньше кандидатов в препараты, отобранных на основании лабораторных и животных исследований, в конечном счёте достигает пациентов. В области психиатрии стресс-связанных расстройств показатели неудач ещё выше, что отражает особую сложность нервной системы и психических процессов по сравнению с другими терапевтическими областями. Десятилетия интенсивных исследований молекулярных механизмов депрессии, тревоги, посттравматического стрессового расстройства генерировали сотни потенциальных мишеней для фармакологического воздействия, но клиническая реализация этих открытий остаётся минимальной. После внедрения селективных ингибиторов обратного захвата серотонина в конце тысяча девятьсот восьмидесятых годов не было создано принципиально новых классов антидепрессантов с существенно улучшенной эффективностью, несмотря на экспоненциальный рост знаний о нейробиологии настроения и стресса.
Конкретные примеры трансляционных неудач в области стресс-связанных расстройств иллюстрируют типичные паттерны разрыва между доклиническими и клиническими результатами. Антагонисты рецепторов кортиколиберина представляли собой теоретически обоснованную стратегию лечения депрессии и тревоги: кортиколиберин является ключевым медиатором стрессового ответа, его рецепторы экспрессируются в областях мозга, связанных с эмоциями, и блокада этих рецепторов у грызунов стабильно снижала тревожное и депрессивноподобное поведение. Клинические испытания нескольких антагонистов кортиколибериновых рецепторов у пациентов с депрессией и тревожными расстройствами, однако, не показали значимого превосходства над плацебо. Аналогичная судьба постигла ингибиторы фосфодиэстеразы, модуляторы нейрокининовых рецепторов, антагонисты глутаматных рецепторов и множество других соединений, которые демонстрировали убедительную эффективность в животных моделях, но не воспроизводили эти эффекты в клинике. Каждая такая неудача представляет не только финансовые потери, но и годы упущенного времени для пациентов, ожидающих более эффективных методов лечения.
Экономические последствия трансляционного разрыва исчисляются миллиардами долларов и создают серьёзное бремя для фармацевтической индустрии, системы здравоохранения и в конечном счёте для общества. Средняя стоимость разработки одного нового лекарственного препарата от начала исследований до выхода на рынок оценивается в диапазоне от одного до двух с половиной миллиардов долларов, и основная часть этих затрат приходится на клинические испытания препаратов, которые в конечном счёте терпят неудачу. Длительность цикла разработки составляет десять-пятнадцать лет, в течение которых капитал заморожен в проектах с неопределённым исходом. Эти экономические реалии привели к тому, что многие крупные фармацевтические компании существенно сократили или полностью свернули свои программы исследований в области психиатрии, сочтя эту область слишком рискованной и непредсказуемой. Парадоксальным образом, именно в тот момент, когда нейронаука накопила беспрецедентное количество знаний о механизмах психических расстройств, индустриальные инвестиции в их трансляцию снижаются. Это создаёт инновационный вакуум, который академические учреждения и государственные агентства пытаются заполнить, но их ресурсы ограничены по сравнению с индустриальными.
Медицинские последствия трансляционного разрыва проявляются в сохраняющемся бремени стресс-связанных расстройств для индивидов и общества. Депрессия остаётся одной из ведущих причин инвалидности в мире, а существующие методы лечения эффективны лишь примерно для половины пациентов, причём многие достигают лишь частичной ремиссии. Значительная доля пациентов с тревожными расстройствами и посттравматическим стрессовым расстройством не отвечает адекватно на доступную терапию или страдает от побочных эффектов, снижающих качество жизни и приверженность лечению. Хронический стресс продолжает вносить вклад в развитие сердечно-сосудистых заболеваний, метаболических нарушений, иммунной дисфункции, и профилактические вмешательства остаются недостаточно эффективными. Каждый год без прогресса в трансляции означает миллионы людей, страдающих от расстройств, для которых более эффективные методы лечения теоретически возможны, но практически недоступны. Этическое измерение трансляционного разрыва состоит в том, что научное сообщество располагает знаниями, которые потенциально могли бы облегчить страдания, но не может эффективно преобразовать эти знания в терапевтические инструменты.
Важно подчеркнуть, что трансляционный разрыв не следует интерпретировать как свидетельство провала фундаментальной науки или бесполезности доклинических исследований. Напротив, он указывает на фундаментальные методологические проблемы в организации трансляционного процесса и на недооценку сложности переноса знаний между биологическими системами и уровнями анализа. Доклинические исследования генерируют ценные знания о механизмах на молекулярном, клеточном и системном уровнях; проблема состоит не в этих знаниях как таковых, а в допущениях о том, как они переносятся на человеческий организм и клиническую ситуацию. Понимание источников трансляционного разрыва открывает возможности для его сокращения через совершенствование методологии на всех этапах исследовательского процесса. Последующие разделы данной темы рассматривают конкретные источники разрыва — ограничения животных моделей, проблему гетерогенности человеческих популяций, методологические различия между уровнями исследований — и стратегии их преодоления.
Связь проблематики трансляционного разрыва с содержанием курса проявляется в нескольких существенных аспектах. Модули, посвящённые биологическим механизмам стресса, представляют знания, большая часть которых получена в доклинических исследованиях и требует осторожности при экстраполяции на людей. Модуль об интервенциях рассматривает методы, прошедшие клиническую валидацию, и обсуждает, почему некоторые теоретически обоснованные подходы не подтвердили эффективности в испытаниях. Модуль о передовых исследованиях включает обсуждение новых стратегий преодоления трансляционного разрыва, таких как использование человеческих органоидов, персонализированная медицина и вычислительные подходы к предсказанию клинической эффективности. Нулевой модуль о научной грамотности закладывает понимание иерархии доказательств, в которой результаты животных исследований занимают более низкую позицию, чем клинические данные, именно в силу трансляционного разрыва. Осознание этой проблематики формирует критическое отношение к доклиническим находкам и реалистичные ожидания относительно их клинической применимости.
Историческая перспектива показывает, что осознание трансляционного разрыва как системной проблемы, а не случайных неудач отдельных проектов, сформировалось относительно недавно, в первое десятилетие двадцать первого века, когда накопленная статистика провалов стала слишком выраженной, чтобы её игнорировать. До этого доминировала оптимистическая линейная модель трансляции, согласно которой фундаментальные открытия естественным образом перетекают в прикладные разработки, а те — в клиническую практику. Эта модель предполагала, что достаточно инвестировать в фундаментальную науку, и практические приложения последуют автоматически. Накопившиеся разочарования заставили пересмотреть эту модель и признать, что трансляция требует специальных усилий, методов и институтов, отличных от тех, которые обеспечивают фундаментальные открытия. Инициативы трансляционной науки, создание специализированных центров и программ, изменение критериев финансирования — всё это отражает институциональный ответ на осознание проблемы. Однако полное преодоление трансляционного разрыва остаётся делом будущего, и текущее поколение исследователей работает в условиях этой неразрешённой проблемы.
Теоретическое осмысление трансляционного разрыва связывает его с более широкими эпистемологическими проблемами, обсуждавшимися в первой теме данного урока. Редукционистская логика доклинических исследований, изолирующих отдельные механизмы и мишени, вступает в противоречие с эмерджентной сложностью человеческого организма и психики. Множественная реализуемость психических расстройств означает, что препарат, эффективный для одного механизма, может не работать для другого механизма, приводящего к внешне сходному синдрому. Нисходящая причинность означает, что психологические и социальные факторы модулируют биологические эффекты способами, не учитываемыми в клеточных и животных моделях. Проблемы многоуровневого моделирования, обсуждавшиеся во второй теме, проявляются в трансляционном контексте как трудность переноса результатов между уровнями анализа. В этом смысле трансляционный разрыв представляет собой практическое следствие тех философских и методологических проблем, которые были рассмотрены ранее, и их понимание необходимо для разработки стратегий преодоления разрыва.
Завершая введение в проблематику трансляционного разрыва, необходимо подчеркнуть конструктивную установку, которую следует занять по отношению к этой проблеме. Признание масштаба разрыва не должно вести к нигилизму относительно возможности трансляции или к обесцениванию доклинических исследований. История медицины знает немало примеров успешной трансляции, когда фундаментальные открытия приводили к революционным терапевтическим достижениям, и нет оснований полагать, что такие успехи невозможны в будущем. Вместе с тем наивный оптимизм линейной модели также неуместен, и каждый трансляционный проект должен осуществляться с осознанием рисков и ограничений. Продуктивная позиция состоит в систематическом анализе источников разрыва, разработке методов их минимизации и реалистичной оценке вероятности успеха на каждом этапе. Последующие разделы этой темы детализируют отдельные источники трансляционных проблем и возможные стратегии их преодоления.
3.2 Ограничения животных моделей стресса
Животные модели составляют основу доклинических исследований стресса и стресс-связанных расстройств, предоставляя возможности для экспериментального изучения механизмов, которые невозможно исследовать непосредственно на людях, однако эти модели имеют фундаментальные ограничения, понимание которых необходимо для адекватной интерпретации результатов и осознанного отношения к трансляционному потенциалу доклинических находок. Использование животных, преимущественно грызунов, в исследованиях стресса обосновывается предположением о консерватизме базовых биологических механизмов стрессового ответа в эволюции: гипоталамо-гипофизарно-надпочечниковая ось, симпатоадреналовая система, воспалительные каскады функционируют по сходным принципам у млекопитающих различных видов. Это предположение имеет значительное эмпирическое подтверждение и позволяет использовать животных для изучения базовой биологии стресса. Однако степень переноса результатов на человека существенно ограничивается межвидовыми различиями в организации нервной системы, уникальностью человеческой психики и когнитивных способностей, а также упрощённостью экспериментальных моделей по сравнению с клинической реальностью. Эти ограничения не обесценивают животные модели, но определяют границы их применимости и требуют осторожности при трансляционных выводах.
Межвидовые различия в организации нервной системы представляют первый и наиболее фундаментальный источник ограничений животных моделей стресса. Мозг грызуна, при всём структурном сходстве с мозгом человека на уровне базовых компонентов, существенно отличается по относительному развитию различных областей, особенно тех, которые критичны для высших когнитивных функций и эмоциональной регуляции. Префронтальная кора, играющая ключевую роль в когнитивном контроле, принятии решений, произвольной регуляции эмоций и стрессовых реакций у человека, у грызунов развита значительно слабее и организована иначе. Области человеческой префронтальной коры, такие как дорсолатеральная префронтальная кора, не имеют прямых гомологов у грызунов, что ставит под вопрос возможность моделирования когнитивных аспектов стрессового ответа. Миндалина, гиппокамп и другие лимбические структуры присутствуют у обоих видов, но их взаимодействие с корковыми регионами и роль в регуляции поведения различаются. Нейрохимические системы, такие как серотонинергическая, также демонстрируют межвидовые различия в распределении рецепторов, плотности иннервации, регуляции синтеза и высвобождения нейромедиаторов.
Проблема моделирования человеческих психических расстройств на животных выходит за рамки нейроанатомических различий и затрагивает саму возможность воспроизведения сложных психопатологических феноменов у нечеловеческих видов. Депрессия у человека включает когнитивные симптомы, такие как негативные автоматические мысли, руминация, чувство безнадёжности и бессмысленности, которые по определению требуют развитых когнитивных способностей и, вероятно, языка. Можно ли говорить о «безнадёжности» у крысы, не обладающей способностью к проспективному мышлению и рефлексии над собственным будущим? Посттравматическое стрессовое расстройство характеризуется навязчивыми воспоминаниями, ночными кошмарами, избеганием триггеров, нарушениями самовосприятия — феноменами, которые трудно операционализировать у животных. Модель «выученной беспомощности», широко используемая для изучения депрессии, воспроизводит некоторые поведенческие аспекты, такие как пассивность и снижение активности, но может ли пассивность крысы, подвергнутой неизбегаемому шоку, считаться эквивалентом экзистенциального отчаяния человека с тяжёлой депрессией? Эти вопросы не риторические; они указывают на принципиальные пределы того, что может быть смоделировано.
Проблема антропоморфизма в интерпретации поведения животных представляет собой методологическую ловушку, в которую легко попасть при работе с моделями стресс-связанных расстройств. Антропоморфизм в данном контексте означает приписывание животным психологических состояний, переживаний и мотиваций, характерных для человека, без достаточных оснований полагать, что животные действительно испытывают нечто подобное. Когда исследователь описывает крысу как «депрессивную» или «тревожную» на основании определённых поведенческих показателей, он использует язык, предполагающий субъективные переживания, доступ к которым у исследователя отсутствует. Крыса демонстрирует сниженную активность в тесте принудительного плавания, но переживает ли она безнадёжность или просто адаптируется к ситуации наиболее энергетически эффективным способом? Мышь избегает открытых участков лабиринта, но испытывает ли она тревогу в человеческом смысле или просто следует инстинктивным программам поведения? Строгий методологический подход требует описания поведения животных в операциональных терминах без психологических атрибуций: «депрессивноподобное поведение», «тревожноподобное поведение», а не «депрессия» и «тревога».
Концепция валидности моделей предоставляет формальный язык для оценки того, насколько животная модель адекватно отражает человеческое расстройство, и различает несколько типов валидности с разными импликациями. Лицевая валидность означает поверхностное сходство между проявлениями модели и симптомами расстройства: если модель воспроизводит некоторые внешние признаки депрессии, такие как снижение активности, ангедония, нарушения сна, она обладает лицевой валидностью. Конструктная валидность относится к тому, воспроизводит ли модель предполагаемые механизмы расстройства: если считается, что депрессия связана с дисрегуляцией серотониновой системы, модель с конструктной валидностью должна демонстрировать аналогичную дисрегуляцию. Предсказательная валидность означает способность модели предсказывать эффективность терапевтических вмешательств у людей: если антидепрессант эффективен в модели и эффективен у пациентов, модель обладает предсказательной валидностью. Критическая проблема состоит в том, что многие модели стресса и депрессии обладают приемлемой лицевой валидностью, но ограниченной или неизвестной предсказательной валидностью, что и объясняет трансляционный разрыв.
Детальный анализ наиболее распространённых моделей депрессии и стресса у грызунов иллюстрирует типичные паттерны валидности и ограничений. Тест принудительного плавания, один из наиболее широко используемых тестов для скрининга антидепрессантов, измеряет время неподвижности грызуна, помещённого в цилиндр с водой; считается, что неподвижность отражает «поведенческое отчаяние», а антидепрессанты снижают неподвижность. Тест обладает предсказательной валидностью для классических антидепрессантов, которые использовались для его разработки, но не всегда предсказывает эффективность новых соединений с иными механизмами действия. Более того, снижение неподвижности может отражать неспецифическое повышение моторной активности, а не антидепрессивный эффект как таковой. Модель хронического непредсказуемого стресса, в которой животные подвергаются разнообразным стрессорам на протяжении недель, воспроизводит некоторые аспекты хронического стресса у людей и индуцирует ангедонию, измеряемую по снижению потребления сладкого раствора. Эта модель имеет лучшую лицевую валидность, но её предсказательная валидность также ограничена, и многие соединения, эффективные в этой модели, не продемонстрировали клинической эффективности.
Проблема экологической валидности стрессоров, используемых в животных моделях, добавляет ещё одно измерение ограничений. Стрессоры, применяемые к лабораторным животным, как правило, являются физическими и непосредственными: электрический шок, иммобилизация, принудительное плавание, социальное поражение. Эти стрессоры резко отличаются от типичных стрессоров современного человека, которые преимущественно психосоциальные, хронические, связанные с когнитивной оценкой и антиципацией: финансовая нестабильность, проблемы в отношениях, профессиональные требования, экзистенциальные тревоги. Физический стрессор вызывает немедленный биологический ответ с последующим восстановлением; психосоциальный стрессор может вызывать хроническую активацию стрессовых систем через руминацию и антиципацию даже при физическом отсутствии угрозы. Модели социального стресса, такие как социальное поражение у грызунов, ближе к человеческой ситуации, но даже они не воспроизводят когнитивную сложность человеческих социальных отношений и связанных с ними переживаний. Разрыв между лабораторными стрессорами и реальными стрессорами человеческой жизни вносит вклад в трансляционные проблемы.
Генетическая и средовая стандартизация лабораторных животных, являющаяся условием экспериментального контроля, одновременно создаёт условия, нетипичные для понимания человеческой вариабельности. Инбредные линии мышей и крыс генетически почти идентичны внутри линии, что устраняет генетическую вариацию и позволяет выявлять эффекты экспериментальных манипуляций на чистом фоне. Стандартизированные условия содержания — клетки определённого размера, контролируемое освещение, стандартный корм, отсутствие естественных стимулов — создают воспроизводимую среду. Однако эти условия диаметрально противоположны человеческой ситуации, характеризующейся генетическим разнообразием, бесконечной вариацией средовых условий, уникальностью жизненных историй. Эффект, стабильно наблюдаемый у генетически идентичных животных в контролируемой среде, может быть замаскирован или модифицирован генетической и средовой вариацией у людей. Это одна из причин, почему эффекты, робастные в животных исследованиях, оказываются слабыми или нестабильными в клинических испытаниях с гетерогенными популяциями пациентов.
Практические следствия понимания ограничений животных моделей для исследовательской практики и интерпретации литературы многообразны и существенны. Исследователь, планирующий использовать животную модель, должен осознанно выбирать модель с учётом её валидности для конкретного исследовательского вопроса, а не по традиции или удобству. Интерпретация результатов должна избегать чрезмерных трансляционных обобщений: обнаружение того, что соединение снижает депрессивноподобное поведение у мышей, не означает, что оно будет эффективным антидепрессантом у людей; это означает лишь, что соединение заслуживает дальнейшего изучения. Критическое чтение литературы требует внимания к типу использованной модели, параметрам стрессорного протокола, измеряемым исходам и их соответствию человеческим феноменам. Разработка новых, более валидных моделей, особенно моделей с лучшей предсказательной валидностью, остаётся важной методологической задачей для поля. Связь с содержанием курса проявляется в том, что биологические механизмы, рассматриваемые в модулях со второго по четвёртый, в значительной мере изучены на животных, и понимание ограничений этих исследований необходимо для адекватной интерпретации представленного материала.
Завершая обсуждение ограничений животных моделей, необходимо подчеркнуть, что эти ограничения не означают бесполезности животных исследований, но требуют их позиционирования в системе доказательств соответствующим образом. Животные модели незаменимы для изучения базовых биологических механизмов, которые консервативны в эволюции; для первичного скрининга потенциальных терапевтических агентов; для исследований, невозможных на людях по этическим причинам. Однако их результаты должны рассматриваться как гипотезы для проверки на людях, а не как доказательства клинической эффективности. Интеграция данных животных исследований с данными из других источников — клеточных моделей с человеческими клетками, исследований у здоровых добровольцев, клинических наблюдений — повышает надёжность трансляционных выводов. Разработка критериев для идентификации находок с высоким трансляционным потенциалом на основе паттернов исторических успехов и неудач представляет собой активную область методологических исследований.
3.3 Проблема гетерогенности человеческих популяций
Если ограничения животных моделей представляют одну сторону трансляционного разрыва, связанную с источником доклинических данных, то гетерогенность человеческих популяций представляет другую сторону, связанную с целью трансляции — пациентами, для которых разрабатываются терапевтические вмешательства. Человечество характеризуется беспрецедентным генетическим разнообразием, бесконечной вариацией жизненных условий, культурных контекстов, индивидуальных историй, и это разнообразие имеет прямые следствия для эффективности терапевтических вмешательств. В контрасте со стандартизированными условиями доклинических исследований, где генетически идентичные животные содержатся в контролируемой среде, клинические испытания и тем более реальная клиническая практика сталкиваются с популяциями, в которых каждый индивид уникален. Эффект, наблюдаемый в среднем по такой гетерогенной группе, может маскировать существенные различия между подгруппами: препарат может быть высокоэффективен для одних пациентов и совершенно неэффективен для других, но при усреднении эти эффекты взаимно нивелируются, создавая видимость умеренной или отсутствующей общей эффективности.
Генетическое разнообразие человеческих популяций влияет на все аспекты стрессового ответа и терапевтической реакции, от фармакокинетики до нейробиологических механизмов. Полиморфизмы генов, кодирующих ферменты метаболизма лекарств, определяют, насколько быстро препарат расщепляется в организме и какие концентрации достигаются в плазме и тканях. Генетические варианты рецепторов, на которые нацелены психотропные препараты, влияют на эффективность связывания и последующей сигнальной трансдукции. Полиморфизмы генов, связанных со стрессовым ответом, таких как ген рецептора глюкокортикоидов или белка FKBP5, регулирующего чувствительность глюкокортикоидного рецептора, создают различия в базовой реактивности на стресс и в ответе на терапевтические вмешательства. Полигенные оценки риска, суммирующие эффекты сотен генетических вариантов, предсказывают вероятность развития стресс-связанных расстройств, но также могут быть связаны с дифференциальным ответом на лечение. Генетическая гетерогенность означает, что «один размер не подходит всем», и терапия, эффективная в среднем по популяции, может быть неоптимальной для многих индивидов.
Эпигенетические различия добавляют ещё один слой гетерогенности, связанный с взаимодействием генома и среды на протяжении индивидуального развития. Эпигенетические модификации — метилирование ДНК, модификации гистонов, экспрессия некодирующих РНК — регулируют активность генов без изменения их последовательности и формируются под влиянием средовых факторов, включая стресс. Два человека с идентичными генотипами могут иметь различные эпигенетические профили, если их жизненные истории различались, и эти эпигенетические различия транслируются в различия в экспрессии генов, нейробиологических функциях, уязвимости к стрессу и ответе на лечение. Ранний стресс, в частности, оставляет долговременные эпигенетические следы, которые программируют системы стресс-ответа и влияют на траектории здоровья на протяжении жизни. Пациенты с историей раннего неблагоприятного опыта могут иметь эпигенетические профили, определяющие иной ответ на терапию, чем пациенты без такой истории, даже при сходной текущей симптоматике. Эпигенетическая гетерогенность, в отличие от генетической, динамична и потенциально обратима, что создаёт как сложности для понимания, так и возможности для вмешательства.
Средовое разнообразие, в котором живут люди, контрастирует со стандартизированными условиями лабораторных исследований и вносит существенный вклад в вариабельность терапевтических ответов. Социально-экономический статус, качество питания, физическая активность, экспозиция загрязняющим веществам, доступ к медицинской помощи, социальная поддержка, культурные практики — все эти факторы варьируют между индивидами и популяциями и модулируют как течение стресс-связанных расстройств, так и ответ на лечение. Пациент, живущий в условиях хронического социально-экономического стресса, может по-иному отвечать на антидепрессант, чем пациент в благополучных условиях, даже если их биологические маркеры депрессии сходны. Культурные различия в понимании психического здоровья, стигме, ожиданиях от лечения влияют на приверженность терапии и субъективную оценку эффектов. Коморбидные соматические заболевания, распространённость которых неравномерна в популяции, изменяют фармакокинетику и фармакодинамику лекарств. Все эти источники средовой гетерогенности действуют одновременно и взаимодействуют с генетическими и эпигенетическими факторами, создавая сложную матрицу индивидуальных различий.
Концепция биотипов стресс-связанных расстройств представляет собой попытку структурировать гетерогенность путём выделения биологически однородных подгрупп внутри диагностически определённых категорий. Согласно этой концепции, диагнозы, такие как «большое депрессивное расстройство» или «генерализованное тревожное расстройство», представляют собой зонтичные категории, объединяющие пациентов с различными биологическими механизмами, которые конвергируют к сходной симптоматике. Выделяются, например, воспалительный биотип депрессии, характеризующийся повышенными маркерами воспаления и предположительно связанный с иммунными механизмами патогенеза; метаболический биотип, связанный с нарушениями энергетического обмена и митохондриальной дисфункцией; нейроэндокринный биотип с преимущественной дисрегуляцией гипоталамо-гипофизарно-надпочечниковой оси; биотип с преобладанием нарушений нейропластичности. Эти биотипы не взаимоисключающи и могут перекрываться, но их выделение позволяет целенаправленно разрабатывать терапии, нацеленные на конкретные механизмы. Противовоспалительные интервенции, например, могут быть эффективны преимущественно для пациентов с воспалительным биотипом, но неэффективны для других биотипов.
Проблема клинических испытаний, не учитывающих гетерогенность, состоит в систематическом искажении оценок терапевтической эффективности. Традиционное рандомизированное контролируемое испытание набирает пациентов, соответствующих диагностическим критериям, случайным образом распределяет их на группу активного лечения и группу плацебо, и оценивает средний эффект лечения по сравнению с плацебо. Если терапия эффективна для подгруппы пациентов с определённым биотипом, но неэффективна для остальных, средний эффект будет разбавлен и может не достичь статистической значимости, даже если для части пациентов терапия высокоэффективна. Именно этот механизм может объяснять многие неудачи клинических испытаний препаратов, имевших убедительную доклиническую поддержку: препарат мог быть разработан для определённого механизма, который присутствует только у части пациентов с данным диагнозом, и его эффект для этой подгруппы размывался при усреднении по всей популяции. Ретроспективный анализ неудачных испытаний иногда выявляет подгруппы с значимым ответом, которые не были идентифицированы проспективно.
Стратегия стратификации представляет собой методологический ответ на проблему гетерогенности и направлена на выделение биологически однородных подгрупп до начала лечения для целенаправленного терапевтического воздействия. Стратификация требует биомаркеров, то есть измеримых биологических показателей, которые предсказывают принадлежность к определённому биотипу и ответ на конкретную терапию. Идеальный биомаркер для стратификации должен быть доступным для рутинного измерения, надёжным и воспроизводимым, достаточно чувствительным и специфичным для дифференциации биотипов, и валидированным как предиктор терапевтического ответа. В области стресс-связанных расстройств разработка таких биомаркеров находится на ранней стадии. С-реактивный белок и другие маркеры воспаления предложены для идентификации пациентов, которые могут ответить на противовоспалительные интервенции. Нейровизуализационные маркеры, такие как паттерны связности мозговых сетей, изучаются как потенциальные предикторы ответа на различные типы антидепрессантов. Генетические полигенные оценки рассматриваются как инструменты персонализации, хотя их предсказательная сила пока ограничена. Переход от исследовательских биомаркеров к клинически применимым инструментам стратификации остаётся нерешённой задачей.
Персонализированная или точная психиатрия представляет собой парадигму, в которой гетерогенность рассматривается не как помеха, а как информация, используемая для оптимизации лечения конкретного пациента. В этой парадигме диагноз дополняется индивидуальным профилем, включающим генетические, эпигенетические, биохимические, нейровизуализационные и клинические данные. На основе этого профиля выбирается терапия, наиболее соответствующая индивидуальным характеристикам, а не стандартное лечение для диагностической категории. Мониторинг биомаркеров в процессе лечения позволяет оценивать ответ и корректировать терапию. Эта парадигма вдохновлена успехами персонализированной медицины в онкологии, где генетическое профилирование опухолей направляет выбор таргетной терапии. Однако в психиатрии реализация персонализированного подхода сталкивается со специфическими трудностями: психические расстройства полигенны и полифакторны; биомаркеры менее валидированы, чем в онкологии; патология локализована в мозге, который менее доступен для биопсии и прямого исследования. Тем не менее персонализированная психиатрия остаётся активной областью исследований и развития.
Связь проблемы гетерогенности с содержанием курса проявляется в нескольких модулях. Модуль о патологии, рассматривающий стресс-связанные расстройства, затрагивает проблему гетерогенности депрессии и тревожных расстройств и обсуждает трансдиагностические подходы. Модуль о биомаркерах и измерении включает обсуждение потенциальных маркеров для стратификации и их текущих ограничений. Модуль об интервенциях рассматривает, почему одни пациенты отвечают на лечение, а другие нет, и какие факторы могут предсказывать ответ. Модуль о передовых исследованиях обсуждает персонализированную психиатрию как перспективное направление и вызовы на пути к её реализации. Модуль о контекстах и популяциях систематически рассматривает источники гетерогенности — возрастные, гендерные, социоэкономические, культурные — и их влияние на стресс и терапевтический ответ. Понимание гетерогенности как фундаментальной характеристики человеческих популяций, а не как артефакта или помехи, представляет собой необходимый элемент зрелого научного мышления о стрессе и его лечении.
Практические следствия проблемы гетерогенности для клинической практики и для проектирования исследований существенны и требуют системных изменений в подходах. Для клинициста это означает осознание того, что неудача стандартного лечения у конкретного пациента не обязательно означает бесперспективность терапии; пациент может относиться к биотипу, для которого эффективны иные подходы. Систематическое измерение потенциальных биомаркеров, мониторинг ответа, готовность к персонализации являются элементами практики, ориентированной на гетерогенность. Для исследователя это означает необходимость стратифицированного дизайна клинических испытаний, в которых гетерогенность контролируется через отбор пациентов по биомаркерам или анализируется как источник информации о механизмах эффективности. Накопление данных о предикторах ответа и формирование баз данных для машинного обучения паттернов гетерогенности представляет собой инвестицию в будущую персонализированную медицину. Преодоление разрыва между обещаниями персонализации и текущей реальностью требует времени, ресурсов и методологических инноваций, но направление движения определено.
Завершая обсуждение гетерогенности человеческих популяций как источника трансляционного разрыва, важно подчеркнуть диалектическую природу проблемы. С одной стороны, гетерогенность создаёт трудности для выявления общих закономерностей и разработки универсально эффективных терапий. С другой стороны, понимание структуры гетерогенности открывает возможности для более точного и эффективного лечения подгрупп пациентов, для которых общие подходы не работают. Переход от модели «один препарат для одного диагноза» к модели «правильный препарат для правильного пациента» концептуально обоснован и технологически всё более возможен по мере развития омиксных технологий, биоинформатики и методов машинного обучения. Трансляционный разрыв в его текущем виде частично отражает применение устаревшей парадигмы, не учитывающей гетерогенность, к реальности, которая принципиально гетерогенна. Преодоление разрыва требует не только улучшения доклинических моделей, но и переосмысления клинических испытаний и практики в направлении персонализации.
3.4 Методологические различия: исследования на клеточных культурах, животных моделях и компьютерное моделирование
Доклинические исследования стресса и стресс-связанных расстройств осуществляются на нескольких методологически различных платформах, каждая из которых обладает специфическими преимуществами и ограничениями, и понимание этих различий необходимо для адекватной интерпретации результатов и оценки их трансляционного потенциала. Традиционно выделяют три основных типа доклинических исследований, обозначаемых латинскими терминами, указывающими на среду проведения эксперимента: исследования на изолированных биологических системах вне организма, исследования на целых живых организмах и компьютерное моделирование. Каждый из этих подходов отвечает на разные вопросы, использует разные методы, генерирует разные типы данных и имеет разное отношение к клинической реальности. Выбор между ними определяется исследовательской задачей, но полноценная трансляционная программа, как правило, требует интеграции данных со всех трёх платформ, поскольку только их комбинация позволяет компенсировать ограничения каждого отдельного подхода. Понимание методологической специфики каждого типа исследований составляет необходимый элемент научной грамотности в области стресса.
Исследования на изолированных биологических системах, проводимые на клеточных культурах, срезах тканей, органоидах или изолированных молекулярных системах, предоставляют максимальные возможности для экспериментального контроля и детального анализа механизмов на молекулярном и клеточном уровнях. В таких экспериментах исследователь может точно контролировать концентрации веществ, воздействующих на биологическую систему, варьировать отдельные параметры при постоянстве остальных, использовать генетические манипуляции для изучения роли конкретных генов и белков, применять высокопроизводительные методы скрининга для тестирования тысяч соединений. Клеточные культуры могут быть получены из различных источников: первичные культуры нейронов или глиальных клеток из мозга животных, иммортализованные клеточные линии, индуцированные плюрипотентные стволовые клетки, дифференцированные в нейроны или другие типы клеток. Органоиды, представляющие собой трёхмерные структуры, выращенные из стволовых клеток и воспроизводящие некоторые аспекты организации тканей, занимают промежуточное положение между простыми клеточными культурами и целыми органами. Срезы мозга сохраняют локальную нейронную архитектуру и позволяют изучать синаптическую передачу и пластичность в относительно интактных нейронных цепях.
Ограничения исследований на изолированных системах связаны прежде всего с потерей организменного контекста, который критичен для понимания интегративных функций, характерных для стрессового ответа. Нейрон в культуре лишён связей с тысячами других нейронов, модулирующих влияний со стороны глиальных клеток, воздействия циркулирующих гормонов и иммунных медиаторов, ритмических изменений, связанных с циркадными циклами. Поведение клетки в таких условиях может существенно отличаться от её функционирования в составе целого организма. Воспалительный ответ микроглии в культуре, например, может не отражать её реакцию в мозге, где микроглия взаимодействует с нейронами, астроцитами, эндотелием сосудов и периферическими иммунными клетками. Кортикостероидный ответ нейронов на добавление синтетического глюкокортикоида в среду не воспроизводит сложную динамику естественного кортизолового ответа с его пульсирующим характером, циркадной ритмикой и контекст-зависимой регуляцией. Более того, условия культивирования сами по себе представляют стресс для клеток: искусственная среда, отсутствие естественной архитектуры, температурные флуктуации при манипуляциях могут активировать стрессовые пути и искажать исходное состояние системы.
Исследования на целых живых организмах, преимущественно грызунах, сохраняют интегративный контекст и позволяют изучать системные взаимодействия, поведение и долгосрочные последствия, недоступные на изолированных препаратах. Животное представляет собой целостную биологическую систему с функционирующими нервной, эндокринной, иммунной системами, с поведенческим репертуаром, с возможностью формирования памяти и обучения. Стрессорное воздействие на животное активирует те же интегративные каскады, которые активируются при стрессе у людей, включая координированную активацию гипоталамо-гипофизарно-надпочечниковой оси, симпатоадреналовой системы, изменения в иммунной функции, поведенческие адаптации. Это делает животные модели незаменимыми для изучения системной биологии стресса и для доклинической оценки терапевтических вмешательств, предполагающих системные эффекты. Кроме того, животные модели позволяют проводить лонгитюдные исследования, отслеживающие развитие патологии во времени, что невозможно в острых экспериментах на клеточных культурах. Возможность манипулировать генетикой животных через создание нокаутных, трансгенных или условно мутантных линий предоставляет мощные инструменты для изучения роли конкретных генов в стрессовых процессах.
Ограничения исследований на животных, рассмотренные подробно в предыдущем разделе, включают межвидовые различия в организации нервной системы и психики, проблемы антропоморфизма в интерпретации поведения, ограниченную предсказательную валидность для человеческих терапевтических исходов. К этим эпистемологическим ограничениям добавляются практические и этические соображения. Исследования на животных затратны в терминах времени, ресурсов и инфраструктуры; содержание виварных, уход за животными, проведение длительных поведенческих экспериментов требуют значительных инвестиций. Этические требования к использованию животных в исследованиях, закреплённые в принципах замены, сокращения и усовершенствования, ограничивают количество и характер экспериментов. Общественное мнение становится всё более критичным к использованию животных в научных целях, что создаёт давление на исследовательское сообщество. Низкая воспроизводимость результатов между лабораториями, связанная с различиями в условиях содержания, протоколах, генетическом фоне животных, подрывает надёжность данных и вносит вклад в трансляционный разрыв.
Компьютерное моделирование, или исследования с использованием вычислительных методов, представляет собой быстро развивающееся направление, которое использует математические модели и симуляции для изучения биологических процессов без непосредственного экспериментирования на живых системах. Этот подход охватывает широкий спектр методов: от молекулярного докинга, предсказывающего связывание лиганда с рецептором, через моделирование сигнальных каскадов и метаболических путей, к нейронным сетевым моделям когнитивных функций и агентному моделированию социальных систем. Преимущества вычислительных методов очевидны: они быстры, относительно дёшевы, не требуют животных и не поднимают этических проблем, позволяют исследовать пространство параметров систематически и полно, могут интегрировать разнородные данные из множества источников. Машинное обучение открывает возможности для выявления паттернов в больших массивах данных, предсказания свойств молекул, классификации биологических образцов, идентификации биомаркеров. Системная биология использует вычислительные методы для моделирования целых биологических систем как сетей взаимодействующих компонентов, схватывая эмерджентные свойства, недоступные редукционистскому анализу.
Фундаментальное ограничение компьютерного моделирования состоит в том, что модель хороша настолько, насколько хороши исходные данные и теоретические предположения, на которых она построена. Если модель основана на неполных или ошибочных данных о структуре белка, параметрах реакций, топологии сетей, её предсказания будут соответственно неполными или ошибочными. Модель является упрощением реальности, и выбор того, что упростить, а что сохранить, определяется теоретическими представлениями исследователя, которые могут быть неадекватны. Проблема переобучения в машинном обучении означает, что модель может превосходно описывать данные, на которых обучалась, но плохо предсказывать новые данные, обнаруживая лишь шум вместо реальных закономерностей. Валидация вычислительных предсказаний требует их экспериментальной проверки на биологических системах, возвращая к необходимости экспериментов на клетках или животных. Таким образом, компьютерное моделирование не заменяет экспериментальные подходы, а дополняет их, направляя эксперименты и интегрируя их результаты.
Конкретный пример, иллюстрирующий взаимодействие трёх уровней доклинических исследований и связанные с ним трансляционные проблемы, может быть построен вокруг разработки противовоспалительной терапии депрессии. На клеточном уровне показано, что добавление провоспалительного цитокина интерлейкина-6 к культуре нейронов гиппокампа снижает их выживаемость и нарушает синаптическую пластичность; блокирование интерлейкина-6 антителами предотвращает эти эффекты. На уровне животных моделей показано, что хронический стресс у мышей повышает уровни интерлейкина-6 в мозге и периферии, и что введение антител против интерлейкина-6 снижает депрессивноподобное поведение в тестах принудительного плавания и потребления сахарозы. Вычислительное моделирование предсказывает, что блокирование интерлейкина-6 должно прервать воспалительный каскад и нормализовать функцию нейронных сетей, связанных с настроением. Все три уровня доказательств конвергируют к заключению, что антитела против интерлейкина-6 должны быть эффективны как антидепрессанты. Однако клинические испытания антител против интерлейкина-6 и его рецептора у пациентов с депрессией показали неоднозначные результаты: некоторые исследования выявили умеренные эффекты у подгруппы пациентов с повышенными исходными маркерами воспаления, другие не обнаружили значимых отличий от плацебо. Этот пример демонстрирует, как убедительные доказательства на всех доклинических уровнях могут не транслироваться в клиническую эффективность.
Причины разрыва между доклиническими и клиническими результатами в приведённом примере множественны и включают факторы, специфичные для каждого уровня исследований. Клеточные эксперименты используют концентрации цитокинов и антител, которые могут быть нефизиологичными и не достижимыми в мозге при системном введении. Животные модели могут не воспроизводить воспалительный профиль, характерный для подгруппы пациентов с депрессией, у которых воспаление является ключевым механизмом. Гетерогенность пациентов в клинических испытаниях размывает эффект, который может быть выражен только у части популяции. Компьютерные модели могут не учитывать компенсаторные механизмы, которые активируются при блокировании одного цитокина и поддерживают воспаление через альтернативные пути. Каждый уровень исследований вносит свои искажения, и их кумулятивный эффект может драматически снижать трансляционный потенциал. Это не означает, что антивоспалительный подход бесперспективен; скорее, это указывает на необходимость более тщательной стратификации пациентов, оптимизации дозирования и выбора мишеней, и разработки комбинированных подходов.
Стратегия триангуляции, предполагающая интеграцию данных с разных уровней доклинических исследований, представляет собой методологический ответ на ограничения каждого отдельного подхода. Согласно этой стратегии, уверенность в механизме или терапевтической мишени повышается, когда данные из клеточных экспериментов, животных моделей и вычислительных предсказаний конвергируют к согласующимся выводам. Расхождение данных между уровнями, напротив, служит сигналом о необходимости дополнительного исследования и осторожности в трансляционных выводах. Триангуляция не гарантирует клинического успеха, но повышает его вероятность и позволяет раньше идентифицировать проблемные кандидаты. Современные программы разработки лекарств всё чаще включают требования подтверждения механизма на нескольких платформах до перехода к клиническим испытаниям. Это удлиняет и удорожает доклиническую фазу, но потенциально экономит ресурсы, предотвращая продвижение неперспективных кандидатов в дорогостоящие клинические испытания.
Технологические инновации размывают границы между традиционными категориями доклинических исследований и создают гибридные подходы с потенциально лучшим трансляционным профилем. Органоиды мозга, выращенные из индуцированных плюрипотентных стволовых клеток человека, сохраняют человеческую биологию, но позволяют экспериментальные манипуляции, невозможные на людях. Микрофлюидные системы, называемые «органами-на-чипе», воспроизводят взаимодействие нескольких типов тканей в миниатюризованном формате с контролируемыми условиями. Гуманизированные мыши, несущие человеческие гены или даже человеческие клетки, сочетают преимущества целого организма с элементами человеческой биологии. Виртуальные пациенты, моделируемые компьютерно на основе данных реальных пациентов, позволяют симулировать ответы на терапию до проведения испытаний. Все эти подходы направлены на сокращение трансляционного разрыва через создание модельных систем, более близких к клинической реальности, чем традиционные клеточные культуры или инбредные мыши.
Связь обсуждаемых методологических различий с содержанием курса проявляется в том, как организовано изложение биологических механизмов стресса в различных модулях. Молекулярные механизмы, рассматриваемые в контексте клеточной сигнализации, нейротрансмиссии, гормональной регуляции, преимущественно изучены с использованием клеточных и молекулярных методов. Системные взаимодействия между нервной, эндокринной и иммунной системами исследованы на животных моделях. Нейровизуализационные данные о мозговых сетях получены как на животных, так и на людях, что создаёт возможность прямого сравнения. Шестой модуль о методах измерения рассматривает различные платформы сбора данных и их ограничения. Одиннадцатый модуль о передовых исследованиях обсуждает новые методологии, включая органоиды, вычислительные подходы и их интеграцию. Понимание того, какие данные получены каким методом и каковы ограничения этого метода, является необходимым условием для критической оценки представленного материала.
3.5 Обратная трансляция: извлечение уроков из клинических неудач
Традиционная модель трансляционного процесса предполагает однонаправленное движение от фундаментальных открытий к клиническим приложениям: базовая наука генерирует знания о механизмах, эти знания воплощаются в терапевтические кандидаты, кандидаты тестируются в клинических испытаниях, успешные препараты внедряются в практику. Эта линейная модель, однако, упускает критически важный элемент: обратную связь от клинических результатов к фундаментальной науке. Обратная трансляция представляет собой стратегию, в которой результаты клинических исследований, особенно неудачных, систематически анализируются для улучшения доклинических моделей, пересмотра теоретических представлений и направления будущих фундаментальных исследований. Когда препарат, убедительно эффективный на животных, не работает у людей, это не просто неудача, подлежащая забвению; это ценная информация о расхождении между моделью и реальностью, которую следует использовать для совершенствования моделей. Систематическое применение обратной трансляции может превратить накопившиеся неудачи трансляционного процесса в источник знаний, направляющих будущие успехи.
Концептуальное обоснование обратной трансляции основывается на признании того, что клинические данные представляют собой наиболее релевантный источник информации о человеческой биологии стресса и его расстройств. Каким бы изощрённым ни был эксперимент на клеточной культуре или животной модели, он изучает модельную систему, а не то, что нас интересует в конечном счёте — человека. Клинические испытания, напротив, проводятся непосредственно на людях с реальными расстройствами и предоставляют данные о том, как терапия работает или не работает в целевой популяции. Когда терапия, основанная на определённой гипотезе о механизме, не достигает клинической эффективности, это ставит под сомнение либо гипотезу, либо способ её проверки на людях, либо модели, на которых гипотеза была разработана. Анализ причин неудачи и возврат этой информации в доклиническую науку создаёт итеративный цикл, в котором каждая неудача приближает к успеху через уточнение понимания и совершенствование методов.
Практическая реализация обратной трансляции требует тщательного анализа причин клинических неудач, что не всегда просто в силу множественности возможных объяснений и часто неполных данных. Когда препарат не показывает эффекта в клиническом испытании, возможные причины включают: неправильную гипотезу о механизме болезни; правильную гипотезу, но неадекватную её реализацию в молекуле препарата; адекватный препарат, но недостаточную экспозицию в мишеневой ткани из-за фармакокинетических проблем; достаточную экспозицию, но неправильный выбор популяции пациентов; правильную популяцию, но неадекватные конечные точки испытания; адекватные конечные точки, но проблемы с дизайном испытания или его проведением. Каждая из этих причин требует разных корректирующих действий и имеет разные импликации для обратной трансляции. Определение истинной причины неудачи часто требует дополнительных исследований, включая анализ биомаркеров, фармакокинетических данных, подгрупповой анализ, механистические исследования в клинических образцах.
Яркий пример обратной трансляции в области нейродегенеративных заболеваний связан с пересмотром амилоидной гипотезы болезни Альцгеймера после серии неудачных клинических испытаний препаратов, нацеленных на бета-амилоид. На протяжении десятилетий доминирующая теория патогенеза болезни Альцгеймера утверждала, что накопление бета-амилоидных бляшек в мозге является первичным патогенетическим событием, и что удаление амилоида должно остановить или замедлить прогрессирование болезни. Многочисленные антиамилоидные препараты, включая моноклональные антитела и ингибиторы ферментов, синтезирующих амилоид, показали впечатляющую эффективность в уменьшении амилоидной нагрузки в мозге, но не улучшали когнитивные функции пациентов. Эти неудачи заставили пересмотреть роль амилоида в патогенезе: возможно, амилоид является маркером, а не причиной нейродегенерации; возможно, вмешательство проводится слишком поздно, когда нейрональное повреждение уже необратимо; возможно, другие патологические процессы, такие как тау-патология или нейровоспаление, более важны для когнитивного снижения. Обратная трансляция этих неудач стимулировала разработку новых доклинических моделей, фокусирующихся на ранних стадиях и альтернативных механизмах, и переориентацию клинических испытаний на более ранние стадии болезни и комбинированные подходы.
В области стресс-связанных расстройств обратная трансляция находит применение в переосмыслении моделей депрессии и тревоги после неудач препаратов, основанных на моноаминергической гипотезе. Моноаминергическая гипотеза, постулирующая дефицит серотонина, норадреналина или дофамина как причину депрессии, доминировала десятилетиями и направляла разработку антидепрессантов. Однако накопившиеся данные указывают на ограниченность этой гипотезы: антидепрессанты повышают уровни моноаминов в синапсе в течение часов, но терапевтический эффект развивается недели; около трети пациентов не отвечают на моноаминергические препараты; многие соединения, мощно модулирующие моноаминовую передачу, не обладают антидепрессивной активностью. Обратная трансляция этих наблюдений привела к смещению фокуса на нижестоящие механизмы, такие как нейропластичность и нейрогенез, на альтернативные системы, такие как глутаматергическая, и на сопутствующие процессы, такие как воспаление. Открытие быстрого антидепрессивного эффекта кетамина, действующего через глутаматную систему, было частично результатом этого переосмысления и само стало источником обратной трансляции, стимулируя исследования механизмов быстрой пластичности.
Институциональные барьеры для эффективной обратной трансляции объясняют, почему эта стратегия реализуется недостаточно систематически, несмотря на её очевидную ценность. Коммуникационный разрыв между доклиническими и клиническими исследователями означает, что информация о клинических неудачах не всегда достигает тех, кто разрабатывает и использует доклинические модели. Академические исследователи, работающие с животными моделями, могут не следить за результатами клинических испытаний в индустрии; клиницисты могут не знать деталей доклинических экспериментов, предшествовавших неудачному препарату. Публикационное смещение усугубляет проблему: неудачные клинические испытания публикуются реже и менее детально, чем успешные, что ограничивает доступность информации для обратной трансляции. Конкурентные и коммерческие соображения в фармацевтической индустрии препятствуют открытому обмену данными о неудачах, которые рассматриваются как конфиденциальная информация. Академическая система вознаграждений стимулирует публикацию новых открытий, а не рефлексивный анализ причин неудач, что направляет усилия исследователей в иное русло.
Систематический подход к обратной трансляции требует институциональных механизмов, обеспечивающих сбор, анализ и распространение информации о клинических неудачах. Регистры клинических испытаний, требующие предварительной регистрации и обязательной публикации результатов независимо от исхода, повышают доступность данных о неудачах. Инициативы по обмену данными между индустрией и академией создают возможности для независимого анализа неудачных испытаний. Специализированные журналы и платформы для публикации негативных результатов снижают публикационное смещение. Междисциплинарные конференции и рабочие группы, объединяющие доклинических и клинических исследователей, способствуют передаче знаний и совместному анализу причин разрыва. Финансирующие агентства могут стимулировать обратную трансляцию, включая её как критерий оценки грантовых заявок и как требуемый компонент трансляционных программ. Все эти механизмы постепенно внедряются, но их распространение остаётся неполным, и культурные изменения в отношении к неудачам происходят медленно.
Методологические следствия обратной трансляции для разработки и использования доклинических моделей существенны и направлены на повышение их предсказательной валидности. Если определённая модель депрессии систематически не предсказывает клиническую эффективность, это сигнал к её пересмотру или замене. Анализ того, какие характеристики модели и препарата коррелируют с клиническим успехом или неудачей, позволяет идентифицировать более предсказательные модели и парадигмы. Переход от простых поведенческих тестов к более комплексным оценкам, включающим когнитивные, социальные и гедонистические домены, может повысить соответствие человеческой симптоматике. Включение биомаркеров, которые можно измерять и у животных, и у людей, создаёт мост для трансляции и позволяет проверять, воспроизводятся ли биомаркерные эффекты при переходе от модели к клинике. Использование разнообразных генетических линий животных вместо стандартных инбредных линий может лучше отражать человеческую вариабельность. Все эти модификации вытекают из уроков, извлечённых через обратную трансляцию.
Роль обратной трансляции в формировании теоретического понимания стресс-связанных расстройств выходит за рамки совершенствования моделей и затрагивает фундаментальные концепции патогенеза. Когда терапия, основанная на определённой теории, не работает, это повод пересмотреть теорию, а не только модель. Неудачи антагонистов рецепторов кортиколиберина ставят вопрос о роли этого пептида в патогенезе депрессии у людей, которая может отличаться от его роли у грызунов. Неудачи простых противовоспалительных подходов указывают на то, что связь воспаления и депрессии более сложна, чем прямая причинность, и что воспаление может быть биотип-специфичным механизмом, а не универсальным. Успехи кетамина, напротив, подтверждают роль глутаматергической системы и быстрой синаптической пластичности в патофизиологии и терапии депрессии. Каждый клинический результат, успешный или нет, представляет собой тест теории на людях и должен использоваться для уточнения теоретических представлений. Наука о стрессе, интегрирующая обратную трансляцию, становится более эмпирически обоснованной и менее зависимой от экстраполяций из модельных систем.
Связь обратной трансляции с содержанием курса проявляется в том, как материал представляет историю развития идей о механизмах стресса и его расстройств. Модуль о нейроэндокринных осях рассматривает эволюцию представлений о роли гипоталамо-гипофизарно-надпочечниковой оси в депрессии, включая переосмысление, вызванное клиническими неудачами. Модуль об интервенциях обсуждает, почему некоторые теоретически обоснованные подходы не подтвердили эффективности и какие уроки из этого извлечены. Модуль о передовых исследованиях рассматривает, как обратная трансляция направляет текущие исследовательские программы. Нулевой модуль о научной грамотности формирует установку на критическую оценку данных и готовность пересматривать представления в свете новых доказательств, что является культурной основой для обратной трансляции. Понимание науки как итеративного процесса, в котором неудачи столь же информативны, как успехи, составляет существенный элемент научного мышления, формируемого курсом.
Перспективы развития обратной трансляции связаны с созданием более систематических и формализованных подходов к извлечению и применению уроков клинических неудач. Базы данных, интегрирующие информацию о доклинических и клинических характеристиках препаратов с их клиническими исходами, позволят применять машинное обучение для выявления паттернов, предсказывающих успех или неудачу. Проспективное планирование обратной трансляции, включающее заранее определённые исследования механизмов в случае неудачи, повысит качество извлекаемой информации. Культурные изменения в академии и индустрии, нормализующие неудачу как источник знаний, а не как повод для стыда, создадут более благоприятную среду для открытого обсуждения и анализа негативных результатов. Всё это направлено на превращение накопленного опыта неудач в стратегический ресурс для будущих успехов, замыкая цикл трансляции и делая его более эффективным.
3.6 Стратегии преодоления трансляционного разрыва: современные инициативы
Осознание масштаба и причин трансляционного разрыва стимулировало разработку разнообразных стратегий его преодоления, направленных на модификацию различных звеньев трансляционной цепи: от совершенствования доклинических моделей до реорганизации клинических испытаний и изменения институциональных структур науки. Эти стратегии не представляют собой единого решения, поскольку такого решения не существует; скорее, они образуют комплекс взаимодополняющих подходов, каждый из которых адресует определённые источники разрыва. Их реализация требует технологических инноваций, методологических разработок, институциональных реформ и культурных изменений в исследовательском сообществе. Некоторые из этих стратегий уже внедряются и демонстрируют обнадёживающие результаты; другие остаются на стадии разработки или сталкиваются с практическими препятствиями. Обзор современных трансляционных инициатив позволяет оценить направление развития поля и перспективы сокращения разрыва в обозримом будущем.
Использование человеческих клеток и органоидов в доклинических исследованиях представляет собой стратегию, направленную на преодоление межвидовых различий, ограничивающих трансляцию результатов с животных моделей. Индуцированные плюрипотентные стволовые клетки, полученные путём перепрограммирования соматических клеток взрослого человека, могут быть дифференцированы в нейроны, астроциты, микроглию и другие типы клеток нервной системы, сохраняя генетический профиль донора. Это позволяет создавать клеточные модели с человеческой биологией, изучать эффекты генетических вариантов, ассоциированных с риском психических расстройств, и тестировать терапевтические кандидаты на клетках человека до перехода к клиническим испытаниям. Церебральные органоиды, представляющие собой трёхмерные структуры, самоорганизующиеся из стволовых клеток и воспроизводящие некоторые аспекты развития и организации мозга, добавляют измерение тканевой архитектуры к клеточным моделям. Органоиды развивают слоистую структуру, напоминающую кортекс, демонстрируют спонтанную нейронную активность, формируют синаптические связи и могут культивироваться месяцами, позволяя изучать долгосрочные процессы. Ограничения органоидов включают отсутствие васкуляризации, ограниченный размер, вариабельность между образцами и неполное воспроизведение зрелой мозговой архитектуры, но технологии быстро развиваются, адресуя эти проблемы.
Гуманизированные животные модели представляют альтернативный подход, сочетающий преимущества целого организма с элементами человеческой биологии. Мыши с человеческими генами, введёнными через трансгенез или замену соответствующих мышиных генов, позволяют изучать функцию человеческих белков в контексте целого организма. Для исследований стресса особенно релевантны гуманизированные модели для генов, связанных со стрессовым ответом и уязвимостью к психическим расстройствам, таких как гены рецепторов глюкокортикоидов, транспортёров моноаминов, нейротрофических факторов. Химерные модели, в которых человеческие клетки, например иммунные клетки или нейроны, пересаживаются в организм мыши, создают системы с человеческими компонентами, функционирующими в живом организме. Мыши с гуманизированной иммунной системой позволяют изучать нейроиммунные взаимодействия с использованием человеческих иммунных клеток. Ограничения гуманизированных моделей связаны с неполнотой гуманизации, возможными конфликтами между человеческими и мышиными компонентами, и сохранением межвидовых различий в негуманизированных системах.
Разработка транслируемых биомаркеров представляет собой стратегию создания измерений, которые могут быть проведены и у животных, и у людей, обеспечивая прямое сравнение и верификацию трансляции. Если биомаркер, отражающий механизм действия препарата, изменяется сходным образом в доклинической модели и у пациентов, это повышает уверенность в том, что механизм транслируется. Нейровизуализационные методы, такие как функциональная магнитно-резонансная томография или позитронно-эмиссионная томография, могут применяться и у грызунов, и у людей с использованием сходных протоколов, позволяя сравнивать паттерны активности мозга и связывание лигандов. Электрофизиологические маркеры, такие как вызванные потенциалы или осцилляторная активность, измеримы у разных видов и могут служить индикаторами нейронной функции. Биохимические маркеры в крови или спинномозговой жидкости, включая гормоны, цитокины, метаболиты, белки, также транслируемы при условии учёта межвидовых различий в референсных значениях и динамике. Поведенческие парадигмы, спроектированные для измерения эквивалентных когнитивных функций у животных и людей, такие как тесты рабочей памяти или внимания, создают трансляционный мост на уровне поведения.
Экспериментальная медицина у людей, или исследования механизмов на ранних фазах клинической разработки, представляет стратегию раннего тестирования гипотез непосредственно на людях до проведения полномасштабных испытаний эффективности. В таких исследованиях небольшие группы здоровых добровольцев или пациентов получают экспериментальное вмешательство, и измеряются биомаркеры, отражающие предполагаемый механизм действия. Если препарат не демонстрирует ожидаемого механистического эффекта на этой стадии, это сигнал к пересмотру дозирования, выбора популяции или даже самой гипотезы, до того как будут затрачены ресурсы на масштабные испытания. Например, антивоспалительный препарат может быть тестирован на способность снижать уровни провоспалительных маркеров в крови и спинномозговой жидкости у здоровых добровольцев после индукции воспаления; только при подтверждении механистического эффекта препарат продвигается к испытаниям у пациентов с депрессией и повышенными воспалительными маркерами. Этот подход требует детального понимания механизма и доступности соответствующих биомаркеров, что не всегда возможно, но там, где применим, он повышает эффективность клинической разработки.
Стратификация пациентов с самого начала процесса разработки представляет парадигматический сдвиг от модели «одно лекарство для одного диагноза» к персонализированному подходу. Вместо разработки препарата для депрессии в целом и последующего разочарования в неоднородной популяции, этот подход предполагает идентификацию конкретного биотипа или механизма, разработку терапии, нацеленной на этот механизм, и тестирование только у пациентов с соответствующим биотипом, определяемым биомаркерами. Пример: если разрабатывается противовоспалительный антидепрессант, включаются только пациенты с повышенными маркерами воспаления, которые предположительно имеют воспалительный механизм депрессии. Это создаёт более гомогенную популяцию, в которой эффект не размывается неотвечающими пациентами с иными механизмами, и повышает вероятность обнаружения истинного эффекта. Ограничения этого подхода включают необходимость валидированных биомаркеров для стратификации, уменьшение потенциального рынка для препарата, сложности регуляторного одобрения для таргетированных терапий. Тем не менее регуляторные агентства всё более поощряют персонализированные подходы, и прецеденты успешных стратифицированных разработок накапливаются.
Институциональные и структурные изменения в организации трансляционной науки адресуют барьеры, связанные с фрагментацией дисциплин, недостаточной коммуникацией между секторами и несоответствием стимулов. Создание трансляционных центров, объединяющих фундаментальных учёных, клиницистов, биоинформатиков, специалистов по регуляторным вопросам, способствует интеграции компетенций и сокращению коммуникационных разрывов. Программы финансирования, специально нацеленные на трансляционные исследования, стимулируют проекты, направленные на преодоление разрыва, а не только на публикацию фундаментальных находок. Партнёрства между академией и индустрией позволяют комбинировать академическую инновационность с индустриальной экспертизой в разработке препаратов. Открытый доступ к данным клинических испытаний и доклинических исследований создаёт ресурсы для мета-анализа, машинного обучения и обратной трансляции. Изменение критериев оценки научной деятельности, включающее не только публикации и цитирования, но и трансляционное влияние, переориентирует стимулы исследователей. Все эти изменения происходят постепенно и неравномерно в разных странах и учреждениях, но их кумулятивный эффект направлен на создание более благоприятной экосистемы для трансляции.
Вычислительные и интегративные подходы используют современные информационные технологии для ускорения и оптимизации трансляционного процесса. Интеграция разнородных данных — омиксных, нейровизуализационных, клинических, литературных — в единые платформы создаёт ресурсы для системного анализа и выявления паттернов, невидимых при фрагментированном рассмотрении. Машинное обучение применяется для предсказания клинической эффективности на основе доклинических характеристик, для идентификации подгрупп пациентов с вероятным ответом на терапию, для репозиционирования существующих препаратов на новые показания. Моделирование виртуальных пациентов и виртуальных клинических испытаний позволяет симулировать исходы до проведения реальных испытаний, оптимизируя дизайн и отбор популяции. Сетевая фармакология анализирует препараты как модуляторы биологических сетей, а не отдельных мишеней, что лучше отражает системную природу заболеваний и терапевтических эффектов. Эти подходы не заменяют экспериментальные и клинические исследования, но повышают их эффективность и направленность, сокращая расточительные итерации методом проб и ошибок.
Регуляторные инновации также вносят вклад в сокращение трансляционного разрыва, адаптируя нормативные рамки к новым научным подходам и ускоряя путь от исследований к практике. Программы ускоренного одобрения для препаратов, адресующих неудовлетворённые медицинские потребности, сокращают время до доступности для пациентов. Адаптивные дизайны клинических испытаний, позволяющие модифицировать протокол в процессе на основе промежуточных данных, повышают эффективность и гибкость. Использование реальных данных и реальных доказательств из электронных медицинских записей, регистров и других источников дополняет данные традиционных клинических испытаний. Регуляторное признание биомаркеров для стратификации и мониторинга создаёт основу для персонализированных подходов. Международная гармонизация регуляторных требований сокращает дублирование и ускоряет глобальный доступ к инновациям. Все эти изменения постепенно трансформируют регуляторный ландшафт в направлении, более благоприятном для трансляции.
Связь стратегий преодоления разрыва с содержанием курса проявляется прежде всего в одиннадцатом модуле о передовых исследованиях, который подробно рассматривает новые методологии и их потенциал для науки о стрессе. Органоиды и человеческие клеточные модели обсуждаются как перспективные платформы для изучения механизмов и скрининга терапий. Мультиомиксная интеграция и вычислительные подходы рассматриваются как инструменты персонализации и предсказания. Институциональные аспекты трансляционной науки затрагиваются в контексте обсуждения будущего поля. Модуль о биомаркерах включает обсуждение транслируемых маркеров и их роли в стратификации. Модуль об интервенциях рассматривает, как современные стратегии разработки влияют на доступность новых терапий. Понимание трансляционного ландшафта и текущих инициатив формирует реалистичные ожидания относительно темпов прогресса и осведомлённость о направлениях развития, актуальную для исследователей, клиницистов и информированных пациентов.
Оценка перспектив преодоления трансляционного разрыва в области стресс-связанных расстройств должна балансировать между осторожным оптимизмом и реалистичным признанием сохраняющихся трудностей. С одной стороны, накопленный опыт неудач, обратная трансляция извлечённых уроков, технологические инновации, институциональные реформы создают предпосылки для более эффективной трансляции в будущем. С другой стороны, фундаментальные проблемы, такие как сложность мозга и психики, гетерогенность расстройств, ограничения моделирования субъективных переживаний, остаются нерешёнными и, возможно, непреодолимыми полностью. Разумные ожидания включают постепенное сокращение разрыва через кумулятивные улучшения на всех этапах, увеличение доли успешных трансляций, появление персонализированных терапий для определённых биотипов, но не революционное исчезновение разрыва в краткосрочной перспективе. Наука о стрессе, как и биомедицина в целом, продвигается инкрементально, через накопление знаний и совершенствование методов, и реалистичное признание этого темпа продвижения предохраняет от нереалистичных обещаний и последующих разочарований.
Завершая обсуждение трансляционного разрыва как темы данного урока, необходимо подчеркнуть её интегрирующее значение для понимания отношений между различными уровнями и типами исследований стресса. Трансляционный разрыв не является изолированной проблемой фармакологической разработки; он отражает фундаментальные эпистемологические трудности переноса знаний между модельными системами и целевой реальностью, между уровнями анализа, между контролируемыми условиями и естественной вариабельностью. Понимание источников разрыва углубляет понимание ограничений каждого типа исследований и каждого уровня анализа, рассмотренных в предшествующих темах. Стратегии преодоления разрыва воплощают принципы интегративного плюрализма, триангуляции и многоуровневого подхода, обсуждавшиеся в философском разделе. Осознание трансляционной проблематики формирует критическое отношение к доклиническим данным, реалистичные ожидания относительно темпов прогресса и понимание того, что путь от открытия к практике долог, неопределёнен и требует систематических усилий по его сокращению.
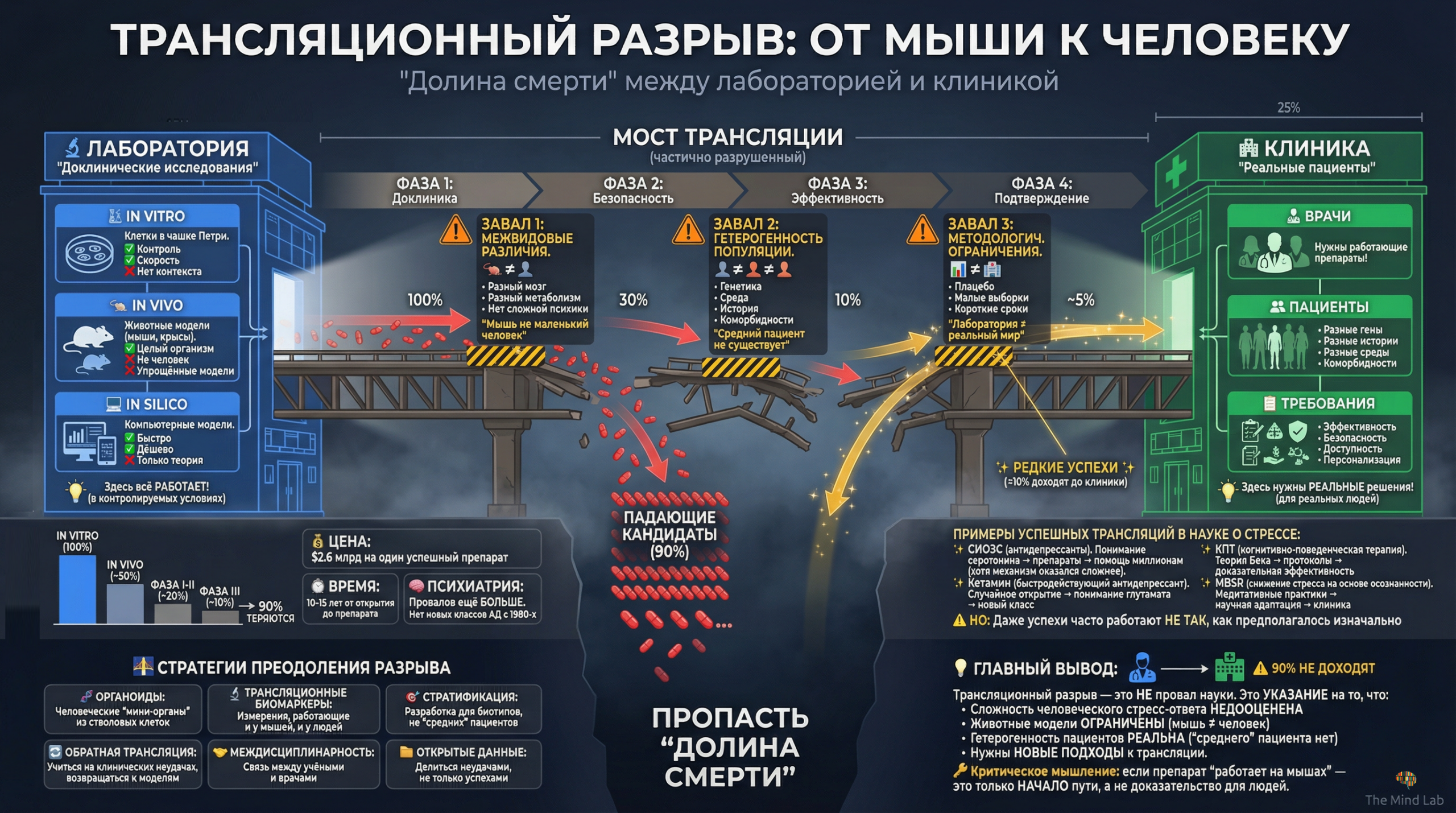
4. Системная биология: сетевой взгляд на стресс
4.1 Парадигмальный сдвиг: от изучения отдельных молекул к анализу сетей взаимодействий
На протяжении большей части истории науки о стрессе доминировал подход, который можно охарактеризовать как молекулоцентричный: исследователи стремились идентифицировать ключевые молекулы, ответственные за стрессовый ответ, и изучали их свойства, функции и регуляцию по отдельности. Кортизол рассматривался как главный гормон стресса, и его эффекты изучались изолированно от других компонентов эндокринной системы. Серотонин исследовался как регулятор настроения, и его недостаточность постулировалась как причина депрессии. Нейротрофический фактор мозга анализировался как молекула пластичности, определяющая способность нейронов к адаптации. Такой подход генерировал обширные знания о свойствах отдельных молекул и принёс значительные успехи, включая разработку фармакологических препаратов, модулирующих конкретные мишени. Однако он также столкнулся с фундаментальными ограничениями, которые становились всё более очевидными по мере накопления данных: изолированное понимание отдельных компонентов не складывалось в целостную картину того, как организм функционирует как интегрированная система, реагирующая на стресс.
Системная биология предлагает радикально иную концептуальную рамку, в которой организм рассматривается не как коллекция отдельных молекул, а как сложная сеть взаимодействующих компонентов, свойства которой определяются структурой и динамикой этих взаимодействий. В этой парадигме отдельная молекула, какой бы важной она ни казалась, не может быть понята вне контекста её связей с другими молекулами; её функция определяется не только её внутренними свойствами, но и её положением в сети, характером её взаимодействий, динамикой этих взаимодействий во времени. Кортизол в этой перспективе — не изолированный агент, а узел в сложной сети, включающей рецепторы, корегуляторы, гены-мишени, метаболические ферменты, транспортёры, связывающие белки, и множество других компонентов, каждый из которых, в свою очередь, связан с другими сетями. Эффект кортизола в конкретной клетке определяется не только его концентрацией, но и состоянием всей сети, в которую он встроен. Этот сдвиг перспективы — от молекул к сетям — представляет собой не просто методологическое усовершенствование, а парадигмальную трансформацию в понимании биологических систем.
Аналогия с социальными системами помогает прояснить логику сетевого подхода и его преимущества перед молекулоцентричным. Понять динамику социальной группы невозможно, изучая индивидов изолированно, какими бы детальными ни были их психологические профили; необходима карта связей между ними, понимание структуры коммуникации, иерархии влияния, паттернов взаимодействия. Влиятельный индивид в центре социальной сети имеет иное воздействие, чем такой же по характеристикам индивид на периферии. Распространение информации, формирование мнений, коллективные решения — всё это эмерджентные свойства сети, не сводимые к свойствам отдельных узлов. Аналогично в биологии: распространение сигнала в клетке, формирование транскрипционного ответа, координация метаболических путей — это сетевые феномены, которые не могут быть поняты через изучение отдельных молекул вне контекста их взаимодействий. Сетевой подход переносит фокус внимания с узлов на связи, со свойств компонентов на топологию их организации, с локальных характеристик на глобальные паттерны.
Концепция эмерджентности, обсуждавшаяся в философском разделе данного урока, находит в сетевом подходе конкретную реализацию и операционализацию. Эмерджентные свойства сетей — это характеристики, которые возникают из взаимодействий компонентов и не присутствуют ни в одном компоненте по отдельности. Синхронизация активности в нейронной сети, например, представляет собой эмерджентный феномен: отдельный нейрон не синхронизирован сам с собой, синхронизация возникает как свойство ансамбля взаимодействующих нейронов. Устойчивость сети к повреждениям — эмерджентное свойство, определяемое топологией связей, а не прочностью отдельных узлов. Способность сети к адаптации и обучению — эмерджентный феномен, возникающий из пластичности связей. В контексте стресса эмерджентность проявляется на многих уровнях: паттерн экспрессии генов в ответ на кортизол — эмерджентное свойство транскрипционной сети; эмоциональное состояние — эмерджентное свойство нейронных сетей мозга; социальная поддержка — эмерджентное свойство сети межличностных отношений. Сетевой подход предоставляет концептуальный и математический аппарат для анализа этих эмерджентных феноменов.
Математический фундамент системной биологии опирается на несколько дисциплин, каждая из которых вносит свои инструменты и концепции в анализ биологических сетей. Теория графов, раздел дискретной математики, предоставляет язык для описания структуры сетей: узлы и рёбра, степень узла, пути и расстояния, кластеризация, централизация. Теория динамических систем, использующая дифференциальные уравнения и анализ фазового пространства, описывает, как состояние сети изменяется во времени под влиянием взаимодействий между компонентами. Теория сложности изучает свойства систем с большим числом взаимодействующих компонентов, нелинейными динамиками и обратными связями. Статистическая механика предоставляет методы для анализа систем с огромным числом степеней свободы через статистические закономерности. Теория информации количественно оценивает потоки информации в сетях. Машинное обучение предлагает алгоритмы для выявления паттернов в высокоразмерных сетевых данных. Интеграция этих математических дисциплин создаёт мощный аналитический аппарат, позволяющий формализовать интуиции о сетевой природе биологических систем и делать количественные предсказания.
Исторический контекст возникновения системной биологии связан с несколькими конвергирующими развитиями в науке и технологии конца двадцатого и начала двадцать первого века. Завершение проекта «Геном человека» предоставило полный каталог генов, но одновременно продемонстрировало, что знание последовательности ДНК недостаточно для понимания функции: необходимо понять, как гены взаимодействуют. Развитие высокопроизводительных технологий — микрочипов для анализа экспрессии, масс-спектрометрии для протеомики, секвенирования следующего поколения — создало возможности для одновременного измерения тысяч молекулярных параметров. Экспоненциальный рост вычислительных мощностей сделал возможным анализ сетей с тысячами узлов и миллионами связей. Концептуальные разработки в области сложных систем, изначально в физике и математике, начали проникать в биологию. Осознание ограничений редукционистского подхода, накопившиеся неудачи трансляции молекулярных открытий в терапию создали запрос на более интегративные подходы. Все эти факторы конвергировали к формированию системной биологии как отдельной дисциплины с собственной методологией и институциональной структурой.
Связь сетевого подхода с содержанием курса о стрессе проявляется в понимании того, как различные компоненты стрессового ответа, рассматриваемые в отдельных модулях, интегрируются в единую систему. Модули о нервной, эндокринной и иммунной регуляции стресса представляют эти системы не как изолированные, а как тесно переплетённые сети с множественными точками взаимодействия. Концепция нейроэндокриноиммунной сети, введённая в первом модуле, получает конкретное воплощение в терминах сетевой топологии: какие узлы связывают эти системы, как сигналы распространяются между ними, как нарушение в одной сети влияет на другие. Модуль о передовых исследованиях детально рассматривает сетевые подходы к моделированию стресса и их применение к пониманию патологии и терапии. Понимание сетевой парадигмы, формируемое в данном разделе, создаёт концептуальную основу для интеграции материала различных модулей в целостную картину.
Практические следствия парадигмального сдвига от молекул к сетям затрагивают все аспекты исследования и терапии стресса. Для исследователя это означает необходимость мыслить в терминах взаимодействий и паттернов, а не отдельных эффекторов; использовать системные методы анализа данных; формулировать гипотезы о сетевых свойствах, а не только о функции отдельных генов или белков. Для клинициста это означает понимание того, что терапия, воздействующая на отдельную мишень, неизбежно затрагивает всю сеть, в которую эта мишень встроена, и что эффекты могут распространяться непредсказуемым образом; что комбинированные подходы, воздействующие на несколько точек сети, могут быть эффективнее монотерапии. Для разработчика препаратов это означает переосмысление стратегий поиска мишеней и дизайна препаратов, рассмотренное детальнее в разделе о сетевой медицине. Сетевой подход — это не просто новый инструмент анализа, а новый способ мышления о биологических системах и их нарушениях.
Завершая введение в сетевую парадигму, важно подчеркнуть, что она не отменяет и не обесценивает молекулярные исследования, а включает их в более широкий контекст. Детальное знание свойств отдельных молекул остаётся необходимым: нельзя понять сеть, не зная свойств её узлов. Однако это знание становится элементом более комплексного понимания, в котором молекула рассматривается не изолированно, а как часть взаимодействующего целого. Редукционистский и системный подходы не противоположны, а комплементарны: редукционизм генерирует знание о компонентах, системный подход интегрирует это знание в понимание целого. Зрелое научное мышление о стрессе требует способности переключаться между этими перспективами, использовать каждую там, где она наиболее продуктивна, и связывать их в когерентную картину. Последующие разделы данной темы конкретизируют сетевой подход применительно к молекулярным сетям, нейронным сетям и динамическим системам, а также рассматривают его терапевтические приложения и ограничения.
4.2 Молекулярные сети: интерактомы и сигнальные пути при стрессе
Концепция интерактома представляет собой операционализацию сетевого подхода на молекулярном уровне и обозначает полную карту всех молекулярных взаимодействий в клетке или организме. Этот термин был введён по аналогии с геномом, обозначающим полный набор генов, и протеомом, обозначающим полный набор белков, и отражает стремление к систематическому картированию взаимодействий как отдельного уровня биологической организации. Интерактом включает различные типы взаимодействий: белок-белковые взаимодействия, в которых два или более белка физически связываются друг с другом; белок-нуклеиновые взаимодействия, в которых белки связываются с ДНК или РНК и регулируют экспрессию генов; метаболические взаимодействия, в которых ферменты катализируют превращения метаболитов; регуляторные взаимодействия, в которых транскрипционные факторы контролируют активность генов. Каждый тип взаимодействий формирует отдельный слой интерактома, и полная картина требует интеграции этих слоёв в многослойную сеть. При стрессе активируются не отдельные изолированные молекулы, а целые модули интерактома, и понимание этих модулей критично для понимания механизмов стрессового ответа.
Экспериментальные методы картирования интерактома развивались на протяжении последних двух десятилетий и достигли масштаба, позволяющего систематически выявлять взаимодействия в геномном масштабе. Дрожжевой двугибридный анализ представляет собой генетический метод, в котором взаимодействие двух белков реконструирует активность транскрипционного фактора и приводит к экспрессии репортёрного гена; этот метод позволяет тестировать миллионы потенциальных белковых пар на взаимодействие. Коиммунопреципитация с последующей масс-спектрометрией использует антитела к интересующему белку для его осаждения вместе со связанными партнёрами, которые затем идентифицируются масс-спектрометрически. Методы бесконтактного мечения позволяют маркировать белки в непосредственной близости к интересующему белку, выявляя пространственные ассоциации. Хроматин-иммунопреципитация с секвенированием картирует места связывания транскрипционных факторов с ДНК в геномном масштабе. Каждый метод имеет свои преимущества и ограничения: двугибридный анализ детектирует прямые бинарные взаимодействия, но вне клеточного контекста; коиммунопреципитация детектирует комплексы в нативных условиях, но не различает прямые и непрямые взаимодействия. Комбинация методов и биоинформатическая интеграция данных позволяют строить всё более полные карты интерактома.
Стрессовый ответ на молекулярном уровне проявляется как координированная активация функциональных модулей интерактома, а не как независимые изменения отдельных молекул. Когда клетка подвергается стрессу, транскрипционный ответ включает изменение экспрессии сотен или тысяч генов, но эти гены не представляют собой случайный набор; они организованы в функциональные кластеры, связанные общими регуляторами и биологическими функциями. Воспалительный модуль включает гены, кодирующие цитокины, хемокины, их рецепторы, сигнальные молекулы, транскрипционные факторы, регулирующие воспаление. Метаболический модуль включает гены ферментов глюконеогенеза, липолиза, аминокислотного катаболизма, обеспечивающие мобилизацию энергетических ресурсов. Модуль защиты от окислительного стресса включает антиоксидантные ферменты, шапероны, компоненты систем репарации ДНК. Эти модули не изолированы, а перекрываются и взаимодействуют: воспалительная сигнализация влияет на метаболизм, метаболические изменения модулируют воспаление. Сетевой анализ позволяет формализовать эти модульные структуры и изучать их взаимодействие.
Концепция сетевых хабов имеет особое значение для понимания молекулярных механизмов стресса и объясняет, почему определённые молекулы играют непропорционально важную роль в регуляции клеточных ответов. Хаб в сетевой терминологии — это узел с необычно большим числом связей, взаимодействующий со многими другими узлами сети. Повреждение или дисфункция хаба имеет непропорционально большие последствия для сети по сравнению с повреждением периферийного узла с малым числом связей, поскольку хаб координирует активность многих компонентов. В молекулярных сетях стресса роль хабов играют так называемые мастер-регуляторы: транскрипционные факторы, регулирующие экспрессию множества генов-мишеней. Ядерный фактор каппа-В представляет собой классический пример воспалительного хаба, регулирующего экспрессию сотен генов, связанных с воспалением, иммунитетом, выживанием клеток. Глюкокортикоидный рецептор является хабом глюкокортикоидного ответа, регулируя тысячи генов-мишеней и взаимодействуя с множеством корегуляторов. Нарушение функции этих хабов катастрофично для соответствующих модулей, что объясняет их центральное положение в патофизиологии стресс-связанных расстройств.
Топологический анализ молекулярных сетей стресса выявляет структурные принципы, определяющие их функцию и уязвимость. Большинство биологических сетей характеризуются безмасштабной топологией, в которой распределение числа связей узлов следует степенному закону: большинство узлов имеет мало связей, но небольшое число хабов имеет очень много связей. Эта топология обеспечивает устойчивость к случайным повреждениям: случайная инактивация узла с высокой вероятностью затронет периферийный узел с малым влиянием на сеть. Однако та же топология делает сеть уязвимой к направленным атакам на хабы: инактивация ключевого хаба может коллапсировать целый модуль. Модульная организация, при которой сеть состоит из относительно автономных функциональных блоков, обеспечивает локализацию повреждений и предотвращает их распространение по всей сети. Иерархическая организация, при которой модули вложены в модули более высокого порядка, обеспечивает координацию функций на нескольких уровнях. Понимание этих топологических принципов имеет прямые терапевтические импликации: воздействие на хабы потенциально более эффективно, но и более рискованно; воздействие на периферийные узлы безопаснее, но менее эффективно.
Динамика молекулярных сетей при стрессе включает не только изменение уровней отдельных компонентов, но и реорганизацию паттернов взаимодействий, что добавляет временное измерение к статической картине топологии. Стресс индуцирует формирование новых белковых комплексов, которые не существовали в нестрессовом состоянии: шапероны связываются с развёрнутыми белками, сигнальные адаптеры рекрутируются к активированным рецепторам, транскрипционные факторы связываются с новыми промоторами. Одновременно другие комплексы диссоциируют: глюкокортикоидный рецептор освобождается от ингибиторного комплекса с шаперонами при связывании кортизола и транслоцируется в ядро. Эти динамические перестройки означают, что интерактом — не статическая карта, а динамическая структура, конфигурация которой зависит от состояния клетки. Временной профиль этих изменений также информативен: ранние изменения могут отражать быстрые посттрансляционные модификации и перераспределение существующих белков, поздние — транскрипционные ответы и синтез новых компонентов. Интегративные подходы, сочетающие статическое картирование взаимодействий с динамическими измерениями, необходимы для полного понимания молекулярных сетей стресса.
Биоинформатические методы играют центральную роль в анализе и интерпретации данных об интерактоме и представляют собой необходимое дополнение к экспериментальным подходам. Экспериментально полученные данные о взаимодействиях интегрируются в базы данных, такие как STRING, BioGRID, IntAct, которые содержат миллионы зарегистрированных взаимодействий и предоставляют инструменты для их анализа. Алгоритмы кластеризации выявляют модульную структуру сетей, группируя тесно связанные узлы. Методы обогащения определяют, какие биологические функции или пути сверхпредставлены среди генов, изменённых при стрессе. Анализ централизации идентифицирует хабы и узлы-мосты, связывающие модули. Методы распространения информации по сети предсказывают, как сигнал, инициированный в одном узле, распространяется по сети. Машинное обучение используется для предсказания неизвестных взаимодействий на основе паттернов известных, для классификации сетевых состояний, для интеграции данных из разных источников. Все эти методы требуют специализированных навыков биоинформатики и становятся всё более необходимыми для современных исследований молекулярных механизмов стресса.
Связь молекулярных сетей с содержанием курса проявляется в модулях, рассматривающих биологические механизмы стресса на молекулярном и клеточном уровнях. Третий модуль о нейроэндокринных осях детально обсуждает сигнальные каскады, инициируемые гормонами стресса, и их интеграцию с другими сигнальными путями; сетевая перспектива помогает понять, как эти каскады переплетаются и координируются. Четвёртый модуль об иммунитете и воспалении рассматривает цитокиновые сети и их взаимодействие с нервной и эндокринной системами; понятие воспалительного модуля как функционально связанного кластера в интерактоме организует этот материал. Шестой модуль о биомаркерах и измерении включает обсуждение омиксных подходов, которые генерируют данные для построения и анализа молекулярных сетей. Одиннадцатый модуль о передовых исследованиях рассматривает системную биологию как одно из ключевых направлений развития науки о стрессе.
Практические следствия понимания молекулярных сетей стресса для исследования и терапии будут детальнее рассмотрены в разделе о сетевой медицине, но заслуживают предварительного упоминания. Идентификация хабов как потенциальных терапевтических мишеней открывает стратегии интервенции с широким спектром эффектов. Понимание модульной организации подсказывает, что воздействие на один модуль может не затрагивать другие, что позволяет более избирательную терапию. Анализ того, какие модули нарушены у конкретного пациента, открывает путь к персонализированному выбору терапии. Предсказание побочных эффектов на основе сетевого распространения — если препарат связывается с мишенью, он влияет на всех её сетевых партнёров — позволяет предвидеть нежелательные реакции. Все эти приложения требуют дальнейшего развития методов и валидации, но концептуально они вытекают из сетевого понимания молекулярной биологии стресса.
4.3 Нейронные сети: коннектомика и функциональная организация мозга при стрессе
Применение сетевого подхода к нервной системе естественно, поскольку мозг представляет собой буквально сеть: миллиарды нейронов, связанных триллионами синапсов, формируют структуру, сетевая природа которой очевидна и на анатомическом, и на функциональном уровне. Коннектомика, наука о картировании нейронных связей, представляет собой применение системно-биологического подхода к нервной системе и стремится к созданию полных карт нейронных связей на различных масштабах: от синаптических связей между индивидуальными нейронами до крупномасштабных трактов, соединяющих области мозга. Различают структурный коннектом, описывающий анатомические связи — аксональные пути и синаптические контакты, и функциональный коннектом, описывающий паттерны совместной активности областей мозга, которые могут отражать функциональную коммуникацию независимо от прямых анатомических связей. При стрессе и стресс-связанных расстройствах обнаруживаются специфические нарушения как структурного, так и функционального коннектома, и их понимание открывает новые перспективы для диагностики и терапии.
Методы картирования структурного коннектома на макроуровне включают прежде всего диффузионную магнитно-резонансную томографию и основанные на ней методы трактографии. Диффузионная томография измеряет направление диффузии молекул воды в ткани мозга, которое ограничено структурой белого вещества: вода диффундирует легче вдоль аксональных пучков, чем поперёк них. На основе этих данных реконструируются траектории нервных трактов, соединяющих различные области мозга, создавая карту структурных путей коммуникации. Современные методы высокоуглового разрешения и вероятностной трактографии позволяют разрешать пересекающиеся волокна и оценивать вероятность существования связей, повышая анатомическую точность реконструкций. Полученные структурные сети описываются матрицами связности, в которых строки и столбцы соответствуют областям мозга, а элементы матрицы отражают силу или вероятность связи между ними. Эти матрицы становятся объектом сетевого анализа с применением методов теории графов: вычисляются метрики централизации областей, кластеризации, эффективности путей, модульной организации. Структурный коннектом предоставляет анатомический каркас, на котором разворачивается функциональная активность мозга.
Функциональный коннектом отражает паттерны совместной активности различных областей мозга и картируется преимущественно с помощью функциональной магнитно-резонансной томографии в состоянии покоя. В этом методе регистрируются спонтанные флуктуации сигнала, связанного с оксигенацией крови, в отсутствие явной задачи, и анализируются корреляции этих флуктуаций между областями мозга. Области, флуктуации активности которых синхронизированы, рассматриваются как функционально связанные, предположительно коммуницирующие и совместно участвующие в когнитивных процессах. Такой анализ выявляет стабильные функциональные сети, воспроизводимые у разных индивидов и в разных исследованиях: сеть пассивного режима работы мозга, активную при самореферентных процессах и отключающуюся при внешне направленном внимании; сеть исполнительного контроля, связанную с рабочей памятью и принятием решений; сеть салиентности, детектирующую значимые стимулы и переключающую другие сети; сенсомоторные и зрительные сети. Функциональная связность между этими сетями и внутри них отражает интеграцию и сегрегацию информационных процессов в мозге и изменяется при психических расстройствах, включая стресс-связанные.
Методы установления каузальных связей в нейронных сетях дополняют корреляционные подходы нейровизуализации и позволяют определить, какие связи являются необходимыми для функции, а какие эпифеноменальны. Оптогенетика, применимая у животных, использует генетически введённые светочувствительные белки для активации или ингибирования специфических популяций нейронов с миллисекундным временным разрешением, позволяя тестировать каузальную роль определённых связей в поведении. Хемогенетика использует модифицированные рецепторы, активируемые только синтетическими лигандами, для модуляции активности определённых нейронов, обеспечивая более длительное, но менее точное во времени вмешательство. У людей каузальные подходы ограничены: транскраниальная магнитная стимуляция позволяет временно модулировать активность поверхностных корковых областей; анализ эффектов локальных повреждений мозга у пациентов предоставляет каузальную информацию о роли отдельных областей. Динамическое каузальное моделирование использует математические модели для инференции направленных связей между областями на основе паттернов активности, хотя результаты зависят от спецификации модели. Комбинация корреляционных и каузальных подходов необходима для полного понимания функциональной архитектуры нейронных сетей.
Изменения функциональной связности мозга при стрессе и стресс-связанных расстройствах демонстрируют специфические паттерны, которые становятся всё более надёжно воспроизводимыми по мере накопления исследований. При хроническом стрессе и депрессии наблюдается гиперсвязность между миндалиной и медиальной префронтальной корой, отражающая усиленную коммуникацию систем детекции угрозы с системами регуляции и самореференции. Одновременно обнаруживается гипосвязность внутри сети пассивного режима работы мозга, которая может отражать дезинтеграцию самореферентных процессов, характерную для депрессии. Сеть исполнительного контроля демонстрирует ослабленную связность с другими сетями, что соответствует когнитивным нарушениям при хроническом стрессе. Баланс между сетью салиентности, которая переключает между внутренне и внешне направленными режимами, и другими сетями нарушается, что может объяснять руминативную застреваемость во внутренне направленном режиме. При посттравматическом стрессовом расстройстве гиперсвязность миндалины особенно выражена и коррелирует с симптомами гипервозбуждения. Важно подчеркнуть, что эти паттерны представляют собой средние тенденции по группам пациентов; индивидуальная вариабельность высока, и ни один паттерн не является диагностическим маркером.
Концепция архитектуры малого мира представляет собой фундаментальный принцип организации мозговых сетей, нарушение которого наблюдается при стрессе и связанных с ним расстройствах. Сети малого мира характеризуются сочетанием высокой локальной кластеризации, при которой соседние узлы тесно связаны между собой, и коротких путей между удалёнными узлами, обеспечиваемых небольшим числом связей-мостов. Эта архитектура оптимально балансирует между локальной специализацией, позволяющей эффективную обработку информации в функциональных модулях, и глобальной интеграцией, позволяющей координацию между модулями. Мозг здорового человека демонстрирует выраженные свойства малого мира как на структурном, так и на функциональном уровне. Хронический стресс и стресс-связанные расстройства ассоциируются с нарушением этого баланса: уменьшается глобальная эффективность сети, отражающая интеграцию, при сохранении или усилении локальной кластеризации; или, наоборот, теряется модульная специализация при усилении хаотических глобальных связей. Степень отклонения сетевой архитектуры от оптимума малого мира коррелирует с тяжестью симптомов и может служить индикатором функционального состояния мозга.
Динамический функциональный коннектом добавляет временное измерение к анализу связности и отражает тот факт, что паттерны функциональной связности не статичны, а изменяются на протяжении времени даже в пределах одного сеанса сканирования. Методы анализа динамической связности используют скользящие временные окна или подходы, не требующие определения окна, для оценки того, как корреляции между областями меняются во времени. Здоровый мозг демонстрирует гибкое переключение между различными конфигурациями связности, соответствующими разным когнитивным режимам или состояниям. При стресс-связанных расстройствах эта динамическая гибкость нарушается: мозг может застревать в одной конфигурации, демонстрировать хаотические переключения без устойчивых состояний или терять способность к адаптивной реконфигурации в ответ на требования среды. Застревание в конфигурации с гиперактивной салиентностью и гипоактивным исполнительным контролем может соответствовать состояниям тревожной руминации; неспособность перейти в режим расслабленного покоя — хронической мобилизации. Динамический анализ обещает более чувствительные маркеры функциональных нарушений, чем статические усреднённые меры связности.
Структурные изменения коннектома при хроническом стрессе отражают долгосрочные последствия стрессовой экспозиции для анатомической организации мозга и связаны с процессами нейропластичности, подробно рассматриваемыми во втором модуле курса. Хронический стресс у животных моделей приводит к ретракции дендритов и потере синапсов в гиппокампе и медиальной префронтальной коре, что транслируется в ослабление структурных связей этих областей. Одновременно наблюдается гипертрофия дендритов в миндалине и орбитофронтальной коре, усиливающая структурную связность систем обработки угрозы и вознаграждения. У людей с хроническим стрессом и стресс-связанными расстройствами обнаруживаются изменения целостности белого вещества, измеряемые показателями диффузионной томографии, в трактах, соединяющих лимбические и префронтальные области. Объём серого вещества редуцирован в гиппокампе и префронтальной коре у пациентов с депрессией и посттравматическим расстройством, хотя причинно-следственные отношения остаются дискуссионными. Эти структурные изменения могут лежать в основе функциональных нарушений связности и определять долгосрочную уязвимость к стрессу.
Переосмысление психических расстройств как сетевых нарушений представляет собой одну из наиболее значимых концептуальных трансформаций в современной психиатрии и непосредственно связано с коннектомным подходом. Традиционная психиатрия мыслила в терминах поломки отдельных структур или дисбаланса отдельных нейромедиаторов: депрессия как дисфункция серотонина, тревога как гиперактивность миндалины, шизофрения как дефицит дофамина. Сетевой подход предлагает альтернативную концептуализацию: психические расстройства представляют собой нарушения паттернов связности и динамики нейронных сетей, а не изолированные поломки отдельных компонентов. Депрессия в этой перспективе — это дисбаланс взаимодействий между сетями эмоциональной обработки, саморегуляции и когнитивного контроля, проявляющийся в специфических паттернах гипер- и гипосвязности. Этот сдвиг имеет диагностические импликации: сетевые метрики могут стать биомаркерами, более чувствительными и специфичными, чем меры активности отдельных областей. Он имеет и терапевтические импликации: интервенции могут быть нацелены на модификацию сетевых паттернов, а не только на отдельные рецепторы или медиаторы.
Связь коннектомного подхода с содержанием курса проявляется прежде всего во втором модуле, посвящённом нервной регуляции и мозгу, который детально рассматривает функции отдельных структур и их взаимодействия. Раздел о мозговых сетях непосредственно применяет коннектомную перспективу к пониманию того, как стресс изменяет паттерны активности и связности ключевых сетей. Модуль о патологии и расстройствах рассматривает сетевые нарушения при конкретных клинических состояниях: посттравматическом расстройстве, депрессии, тревожных расстройствах. Модуль об интервенциях включает обсуждение того, как различные терапевтические подходы модифицируют сетевую организацию мозга: психотерапия через нисходящую регуляцию, фармакотерапия через модуляцию нейромедиаторных систем, нейростимуляция через прямое воздействие на узлы и связи. Модуль о передовых исследованиях рассматривает перспективы использования коннектомных данных для персонализации терапии и предсказания её эффективности.
Методологические ограничения коннектомных исследований заслуживают упоминания для формирования сбалансированной оценки этого подхода. Функциональная связность, измеряемая корреляциями сигнала фМРТ, не эквивалентна анатомическим связям и не обязательно отражает нейронную коммуникацию: области могут коактивироваться из-за общего входа или синхронизирующего воздействия третьей области. Временное разрешение фМРТ порядка секунд не позволяет схватывать быстрые нейронные процессы, происходящие на миллисекундных масштабах. Пространственное разрешение ограничено: стандартный воксел содержит миллионы нейронов, и усреднённый сигнал может маскировать разнородные активности. Парцелляция мозга на области, используемая для построения сетей, в значительной мере произвольна и влияет на результаты анализа. Воспроизводимость индивидуальных коннектомных профилей между сессиями ограничена, что создаёт трудности для клинических приложений. Все эти ограничения не обесценивают коннектомный подход, но требуют осторожности в интерпретации и осознания того, что сетевые карты мозга представляют собой модели реальности, а не саму реальность.
Завершая обсуждение нейронных сетей и коннектомики, важно подчеркнуть интегрирующий потенциал этого подхода для понимания стресса на системном уровне нервной организации. Стресс изменяет мозг не только на уровне отдельных молекул, клеток или структур, но и на уровне организации взаимодействий между структурами, паттернов связности, динамики переключения между режимами функционирования. Коннектомная перспектива схватывает этот уровень организации и предоставляет концептуальный и методологический аппарат для его изучения. Связывая молекулярные и клеточные изменения, рассматриваемые в предшествующих модулях курса, с функциональными и клиническими проявлениями, рассматриваемыми в последующих модулях, коннектомный уровень занимает центральное положение в многоуровневом понимании стресса. Развитие коннектомных методов и накопление данных о сетевых нарушениях при стресс-связанных расстройствах представляет одно из наиболее динамичных направлений современной нейронауки стресса.
4.4 Теория сложных систем: аттракторы, критические переходы и устойчивость
Теория сложных систем предоставляет математический и концептуальный аппарат для понимания динамики биологических систем, выходящий за рамки статического описания сетевой топологии и схватывающий то, как системы ведут себя во времени, как они переходят между состояниями и как они реагируют на возмущения. Организм под стрессом представляет собой парадигматический пример сложной динамической системы: множество взаимодействующих компонентов на разных уровнях организации, нелинейные связи между ними, петли обратной связи, задержки, пороговые эффекты, эмерджентные свойства. Линейные модели, предполагающие пропорциональность между причиной и следствием, неадекватны для описания таких систем: малые изменения могут вызывать катастрофические последствия, а большие воздействия иногда поглощаются без видимых эффектов. Ключевые концепции теории сложных систем — аттракторы, критические переходы, устойчивость, предвестники перехода — предоставляют язык для обсуждения этих нелинейных феноменов и имеют прямые приложения к пониманию стресса, адаптации и развития патологии.
Концепция аттрактора описывает устойчивые состояния, к которым динамическая система стремится при различных начальных условиях и вокруг которых она колеблется при возмущениях. В терминах фазового пространства, где каждая точка представляет конфигурацию всех переменных системы, аттрактор — это область или траектория, к которой сходятся близлежащие траектории. Простейший тип аттрактора — точечный аттрактор, представляющий стационарное состояние: система, выведенная из равновесия, возвращается к нему. Периодический аттрактор, или предельный цикл, представляет устойчивые колебания, как циркадные ритмы физиологических параметров. Странный аттрактор характеризует хаотическую динамику, в которой траектории чувствительны к начальным условиям, но остаются в ограниченной области пространства. Применительно к стрессу и здоровью здоровое состояние может концептуализироваться как аттрактор: система флуктуирует вокруг него, подвергается возмущениям, но возвращается к нему благодаря гомеостатическим механизмам. Болезненное состояние — альтернативный аттрактор, к которому система может перейти при определённых условиях и в котором она затем застревает.
Метафора ландшафта потенциальной энергии помогает визуализировать динамику системы с множественными аттракторами и её поведение под стрессом. Представим ландшафт с холмами и долинами, где долины соответствуют аттракторам, а положение шарика на ландшафте — текущему состоянию системы. Шарик в долине стремится оставаться там; возмущения временно выводят его из низшей точки, но он скатывается обратно. Глубина долины соответствует устойчивости аттрактора: глубокая долина означает высокую устойчивость, требуется большое возмущение для выведения из неё. Высота перевала между долинами определяет барьер перехода между аттракторами: низкий перевал означает лёгкость перехода. Хронический стресс в этой метафоре постепенно изменяет форму ландшафта: долина здоровья становится мельче, долина болезни глубже, перевал понижается. В определённый момент барьер исчезает, и система внезапно переходит в альтернативный аттрактор — происходит срыв. Эта метафора объясняет, почему постепенное накопление стресса может длительно не проявляться клинически, а затем вызвать резкий переход к патологическому состоянию.
Критические переходы, или точки бифуркации, представляют собой моменты качественного изменения динамики системы при постепенном изменении параметров и объясняют внезапность срывов, которая интуитивно кажется несовместимой с постепенностью накопления стресса. В математических терминах бифуркация происходит, когда при изменении управляющего параметра топология фазового пространства качественно меняется: аттрактор может исчезнуть, разделиться, изменить характер. Седло-узловая бифуркация, наиболее релевантная для понимания стрессовых срывов, характеризуется слиянием и взаимным уничтожением устойчивого и неустойчивого равновесий: устойчивое состояние исчезает, и система скачком переходит к альтернативному аттрактору. В контексте стресса это соответствует сценарию, в котором накопление аллостатической нагрузки постепенно ослабляет устойчивость здорового состояния, пока оно не становится неустойчивым и система не переключается в патологический режим. Важно, что этот переход может произойти внезапно, без предупреждения, даже при малом дополнительном стрессоре, который становится «последней каплей».
Устойчивость, или резилентность, в терминах теории сложных систем определяется как способность системы поглощать возмущения, сохраняя при этом свою структуру, функцию и идентичность, то есть оставаясь в текущем аттракторе. Это понятие отличается от обыденного понимания устойчивости как сопротивляемости изменениям и от инженерного понимания как скорости возврата к равновесию. Экологическая устойчивость, концепция, заимствованная из экологии и применимая к биологическим и психологическим системам, характеризует величину возмущения, которую система может абсорбировать без перехода в альтернативный аттрактор. Система может быть устойчива в смысле быстрого возврата к равновесию при малых возмущениях, но неустойчива к большим возмущениям, превышающим порог. Хронический стресс снижает устойчивость, уменьшая бассейн притяжения здорового аттрактора и приближая систему к границе, за которой малое дополнительное возмущение вызовет переход. Понимание устойчивости в этих терминах трансформирует цели интервенции: не только устранение стрессора или симптома, но и восстановление устойчивости системы, расширение её способности абсорбировать будущие возмущения.
Предвестники критического перехода представляют собой особенно интригующий и практически значимый аспект теории сложных систем, открывающий возможности для предсказания и превенции срывов. Математическая теория предсказывает, что вблизи точки бифуркации динамика системы демонстрирует характерные статистические свойства: увеличивается автокорреляция временных рядов — система медленнее возвращается к равновесию после возмущений; увеличивается вариантность — флуктуации становятся более выраженными; возникает мерцание — система начинает колебаться между состояниями. Эти феномены, известные как критическое замедление, отражают ослабление восстанавливающих сил вблизи критической точки и теоретически могут служить ранними сигналами приближающегося перехода. Эмпирические исследования в экологии, климатологии, финансах продемонстрировали, что такие сигналы действительно предшествуют критическим переходам. Применительно к стрессу и психопатологии исследуется, могут ли увеличение вариабельности настроения, замедление восстановления после негативных событий, мерцание между нормальным и субклиническим состояниями служить предвестниками депрессивного эпизода или срыва.
Математический формализм теории динамических систем включает аппарат дифференциальных уравнений, фазовых портретов, анализа устойчивости, бифуркационных диаграмм, который позволяет формализовать и количественно анализировать обсуждаемые концепции. Состояние системы описывается вектором переменных, эволюция во времени — системой дифференциальных уравнений, определяющих, как скорость изменения каждой переменной зависит от текущих значений всех переменных. Фазовое пространство, в котором каждая точка соответствует одной конфигурации переменных, позволяет визуализировать траектории системы и топологию аттракторов. Линеаризация уравнений вблизи равновесий и анализ собственных значений якобиана определяют устойчивость равновесий. Численные методы позволяют симулировать динамику систем, которые не поддаются аналитическому решению. Применение этого формализма к биологии стресса требует идентификации ключевых переменных, построения уравнений на основе биологических знаний, оценки параметров из экспериментальных данных, валидации предсказаний модели. Это технически сложная задача, но она открывает путь к количественным предсказаниям о динамике стресса и эффектах интервенций.
Применение теории сложных систем к пониманию депрессии и других стресс-связанных расстройств генерирует новые гипотезы о природе этих состояний и механизмах их развития. Депрессия может концептуализироваться как альтернативный устойчивый аттрактор: система, попавшая в этот аттрактор, стремится оставаться в нём даже после устранения первоначального стрессора, что объясняет персистенцию депрессивных эпизодов. Рецидивирующий характер депрессии может отражать бистабильность: система способна находиться в двух устойчивых состояниях, и переключения между ними происходят стохастически или под влиянием триггеров. Индивидуальные различия в уязвимости могут отражать различия в форме ландшафта аттракторов: у одних индивидов глубокая долина здоровья и высокий барьер, у других — мелкая долина и низкий барьер. Эффекты раннего стресса могут перманентно изменять ландшафт, создавая предрасположенность. Терапия может работать через расширение бассейна притяжения здорового аттрактора или повышение барьера перехода. Все эти гипотезы, вытекающие из системной перспективы, дополняют традиционные механистические объяснения и открывают новые исследовательские направления.
Практические приложения теории сложных систем к профилактике и терапии стресс-связанных расстройств связаны прежде всего с возможностью предсказания критических переходов и своевременного вмешательства. Если предвестники перехода действительно детектируемы, возникает возможность идентифицировать индивидов, приближающихся к срыву, до того как срыв произойдёт, и вмешаться превентивно. Методы интенсивного лонгитюдного мониторинга с помощью носимых устройств и экологической моментальной оценки создают потоки данных, в которых можно искать статистические сигналы критического замедления: увеличение автокорреляции, нарастание вариабельности, удлинение времени восстановления. Алгоритмы машинного обучения могут обучаться распознавать эти паттерны и генерировать предупреждения. Интервенции, направленные на повышение устойчивости — укрепление копинговых ресурсов, социальной поддержки, физиологической регуляции — могут отодвигать критическую точку и предотвращать переход. Эта парадигма превентивной психиатрии, основанной на предсказании и упреждающем вмешательстве, находится на ранней стадии развития, но представляет одно из перспективных направлений трансляции теории сложных систем в клиническую практику.
Связь теории сложных систем с содержанием курса наиболее непосредственно проявляется в одиннадцатом модуле о передовых исследованиях, который детально рассматривает сетевые и динамические модели стресса как одно из ключевых направлений развития поля. Концепция аллостатической нагрузки, центральная для первого модуля, получает динамическую интерпретацию в терминах постепенного ослабления устойчивости и приближения к критическому переходу. Понимание резилентности, обсуждаемое в контексте протективных факторов в модуле о патологии и о контекстах, обогащается формальным определением устойчивости как свойства динамической системы. Модуль об интервенциях может рассматривать терапевтические эффекты в терминах модификации динамики системы и ландшафта аттракторов. Шестой модуль о методах измерения включает обсуждение интенсивных лонгитюдных подходов, генерирующих временные ряды для анализа динамики. Теория сложных систем, таким образом, предоставляет интегративный концептуальный каркас, связывающий различные аспекты курса в когерентную динамическую картину.
Завершая обсуждение теории сложных систем, важно отметить, что эта перспектива дополняет, а не заменяет молекулярные, клеточные и системные описания стресса. Знание аттракторов и бифуркаций не освобождает от необходимости понимать конкретные биологические механизмы, реализующие эту динамику: какие молекулы, какие клетки, какие нейронные цепи. Динамическая перспектива отвечает на вопросы «как система ведёт себя во времени» и «почему переход происходит внезапно», но не на вопрос «через какие механизмы». Интеграция динамического и механистического понимания представляет собой важнейшую задачу для зрелой науки о стрессе: динамические модели должны быть обоснованы механистически, а механистические знания должны быть интегрированы в понимание системной динамики. Эта интеграция составляет одну из целей курса в целом и находит отражение в связывании материала различных модулей.
4.5 Сетевая медицина: новые подходы к терапевтическим интервенциям
Сетевая медицина представляет собой трансляционное применение системной биологии к клинической практике и основывается на переосмыслении болезней как возмущений в молекулярных и клеточных сетях, а терапии как вмешательств, направленных на восстановление нормальной сетевой функции. Традиционная парадигма «одна болезнь — одна мишень — один препарат» предполагала, что каждое заболевание вызвано нарушением конкретного гена, белка или пути, и что блокирование или активация этой мишени устранит проблему. Эта парадигма принесла успехи в некоторых областях, особенно при инфекционных заболеваниях и моногенных расстройствах, но оказалась недостаточной для сложных мультифакториальных заболеваний, включая стресс-связанные расстройства. Сетевая медицина предлагает альтернативную логику: болезни затрагивают модули сетей, а не отдельные компоненты, и эффективная терапия должна модулировать сетевые паттерны, а не изолированные мишени. Это открывает новые стратегии разработки и применения терапевтических вмешательств, включая полифармакологию, комбинированную терапию и репозиционирование препаратов.
Полифармакология как терапевтическая стратегия основывается на целенаправленном использовании препаратов, действующих на несколько молекулярных мишеней одновременно, в противоположность традиционному стремлению к максимальной селективности. Классические психотропные препараты, включая трициклические антидепрессанты и типичные антипсихотики, являются полифармакологическими по своей природе: они взаимодействуют с множеством рецепторов, транспортёров, ферментов. Это традиционно рассматривалось как недостаток, источник побочных эффектов, и усилия направлялись на создание более селективных соединений. Однако сетевая перспектива предлагает альтернативную интерпретацию: полифармакологическое воздействие на несколько узлов сети может быть более эффективным для модуляции сетевой динамики, чем высокоселективное воздействие на один узел, которое компенсируется перестройкой сети. Неудачи многих высокоселективных препаратов в клинических испытаниях, обсуждавшиеся в разделе о трансляционном разрыве, могут частично объясняться этим механизмом. Целенаправленный дизайн полифармакологических препаратов с оптимизированным профилем связывания нескольких мишеней представляет перспективное направление сетевой фармакологии.
Комбинированная терапия представляет собой стратегию одновременного применения нескольких препаратов, каждый из которых воздействует на разные узлы или модули патогенетической сети. В отличие от монотерапии, стремящейся к максимальному эффекту через одну мишень, комбинированный подход использует синергию между несколькими умеренными эффектами для достижения общего результата. Каждый препарат в комбинации может применяться в более низкой дозе, чем при монотерапии, что снижает риск дозозависимых побочных эффектов. Воздействие на несколько точек сети затрудняет компенсаторную адаптацию, которая ограничивает эффективность монотерапии. В онкологии комбинированная терапия давно стала стандартом, и её успехи послужили моделью для других областей. В психиатрии стресс-связанных расстройств комбинации антидепрессантов с разными механизмами, антидепрессантов с антипсихотиками или анксиолитиками применяются эмпирически, но сетевой подход предлагает рациональную основу для выбора комбинаций на основе анализа того, какие модули сети нарушены у конкретного пациента и какие препараты их модулируют.
Репозиционирование препаратов использует сетевой анализ для идентификации новых терапевтических применений существующих лекарств, что существенно сокращает время и стоимость разработки по сравнению с созданием новых молекул. Если препарат, одобренный для одного показания, модулирует сетевой модуль, вовлечённый в патогенез другого заболевания, он может быть эффективен при этом заболевании. Сетевой анализ интерактомов болезней, в которых картируются гены и белки, ассоциированные с каждым заболеванием, позволяет выявлять перекрытия между болезнями и идентифицировать препараты, мишени которых находятся в зоне перекрытия. Примером, релевантным для стресс-связанных расстройств, является метформин: препарат первой линии для диабета второго типа, который, согласно сетевому анализу, воздействует на пути, связанные с воспалением, митохондриальной функцией, метаболизмом глюкозы в мозге — все эти пути вовлечены в патофизиологию депрессии. Эпидемиологические данные и предварительные клинические исследования поддерживают потенциальную эффективность метформина при депрессии, хотя крупные контролируемые испытания ещё не завершены. Аналогичная логика применяется к статинам, противовоспалительным препаратам, модуляторам инсулиновой сигнализации.
Персонализированный выбор терапии на основе сетевого профиля пациента представляет собой долгосрочную цель сетевой медицины, связывающую её с концепцией биотипов, обсуждавшейся ранее. Если у разных пациентов с одним диагнозом нарушены разные сетевые модули, то оптимальная терапия для каждого должна быть нацелена на его специфический профиль нарушений. Пациент с преобладанием воспалительного модуля может отвечать на противовоспалительные интервенции; пациент с преобладанием митохондриальной дисфункции — на метаболические модуляторы; пациент с дисбалансом нейротрансмиттерных систем — на классические психотропные препараты. Реализация этой стратегии требует доступных и валидированных методов характеризации сетевого профиля у индивидуального пациента: омиксного профилирования, биомаркерных панелей, нейровизуализационных сигнатур. Она также требует понимания того, какие терапевтические воздействия модулируют какие сетевые модули, что пока изучено недостаточно. Несмотря на эти ограничения, концептуальный каркас персонализированной сетевой медицины определяет направление развития, и отдельные элементы, такие как выбор противовоспалительной терапии для пациентов с высоким С-реактивным белком, уже исследуются в клинических испытаниях.
Сетевое предсказание побочных эффектов представляет практически важное приложение сетевой фармакологии, использующее понимание того, что препарат, связываясь с мишенью, влияет на всех её сетевых партнёров. Традиционная токсикология оценивает побочные эффекты эмпирически в клинических испытаниях и постмаркетинговом наблюдении. Сетевой подход позволяет предсказывать некоторые побочные эффекты до клинических испытаний на основе анализа того, какие биологические процессы связаны с мишенями препарата через сеть взаимодействий. Если препарат связывается с рецептором, который через несколько шагов связан с кардиотоксическими путями, это сигнал для дополнительного мониторинга сердечных эффектов. Такое предсказание никогда не будет абсолютно надёжным, поскольку сетевые модели неполны, а эффекты зависят от контекста, но оно может направлять доклиническую и клиническую оценку безопасности и идентифицировать неочевидные риски.
Интеграция сетевой медицины в клиническую практику требует преодоления ряда барьеров, включающих технологические, регуляторные и образовательные компоненты. Технологически необходимы доступные и надёжные методы характеризации сетевых профилей пациентов, которые могут быть использованы в рутинной клинической практике, а не только в исследовательских условиях. Биоинформатическая инфраструктура для анализа и интерпретации сетевых данных должна стать частью клинических информационных систем. Регуляторные рамки, разработанные для оценки препаратов по парадигме «одна мишень — один эффект», требуют адаптации для полифармакологических и комбинированных подходов. Образование клиницистов должно включать основы системного мышления, сетевой биологии, интерпретации многомерных данных. Все эти изменения происходят постепенно, и внедрение сетевой медицины в рутинную практику — процесс, измеряемый десятилетиями, а не годами. Однако концептуальный сдвиг уже происходит, и понимание его направления важно для исследователей и клиницистов, формирующих будущее поля.
Связь сетевой медицины с содержанием курса проявляется прежде всего в модуле об интервенциях, который рассматривает различные терапевтические подходы и их механизмы. Понимание терапии как модуляции сетей, а не воздействия на изолированные мишени, обогащает интерпретацию того, как работают фармакологические и нефармакологические интервенции. Психотерапия в сетевой перспективе может рассматриваться как модуляция нейронных сетей через нисходящие когнитивные процессы; социальная поддержка — как буферирование стрессовых воздействий через социальные сети; изменение образа жизни — как комплексная модуляция метаболических, воспалительных, нейроэндокринных модулей. Модуль о передовых исследованиях детально рассматривает перспективы сетевой медицины и вычислительной психиатрии. Модуль о биомаркерах обсуждает методы характеризации, необходимые для реализации персонализированного сетевого подхода.
4.6 Ограничения и критика системной биологии
При всём потенциале системной биологии для понимания и терапии стресса этот подход сталкивается с существенными ограничениями, которые необходимо осознавать для формирования сбалансированной оценки и предотвращения нереалистичных ожиданий. Критический анализ ограничений не обесценивает системный подход, но определяет условия его продуктивного применения и области, в которых традиционные редукционистские методы остаются незаменимыми. Системная биология — молодая дисциплина, находящаяся в стадии активного развития, и многие её методы ещё не достигли зрелости, необходимой для надёжного клинического применения. Понимание текущих ограничений помогает направлять будущие методологические разработки и предохраняет от преждевременных и необоснованных трансляционных обещаний.
Проблема вычислительной сложности составляет первое фундаментальное ограничение системно-биологического анализа. Биологические сети содержат тысячи и десятки тысяч узлов и сотни тысяч или миллионы связей между ними. Полный анализ динамики таких сетей, учитывающий нелинейности, петли обратной связи, временные задержки, стохастичность, выходит за пределы современных вычислительных возможностей даже при использовании суперкомпьютеров. Симуляции полных молекулярных моделей клетки остаются недоступными, и анализ вынужденно ограничивается подсетями, агрегированными представлениями, упрощёнными моделями динамики. Методы снижения размерности, выделения ключевых компонентов, иерархического моделирования позволяют частично справляться с этой сложностью, но неизбежно вносят упрощения, последствия которых не всегда предсказуемы. На уровне мозговых сетей проблема ещё острее: полный коннектом человеческого мозга с синаптическим разрешением содержал бы порядка сотни триллионов связей, что превышает возможности хранения и анализа. Работа с агрегированными представлениями на уровне областей мозга теряет информацию о микроструктуре сетей.
Неполнота и ненадёжность данных о биологических сетях представляет второе критическое ограничение, ставящее под вопрос валидность сетевых моделей, построенных на этих данных. Экспериментальное картирование интерактомов далеко от завершения: по различным оценкам, известна лишь небольшая доля всех белок-белковых взаимодействий в клетке человека. Более того, известные взаимодействия загрязнены ложноположительными результатами, возникающими из-за технических артефактов экспериментальных методов, и ложноотрицательными пробелами, когда реальные взаимодействия не детектируются. Разные экспериментальные методы выявляют разные наборы взаимодействий с ограниченным перекрытием, что указывает на методическую зависимость результатов. Данные из баз интерактомов представляют собой гетерогенную смесь результатов из разных источников, разного качества, полученных в разных экспериментальных условиях. Сетевой анализ, основанный на таких неполных и зашумлённых данных, может приводить к выводам, отражающим артефакты данных, а не биологическую реальность.
Проблема переобучения и недостаточной генерализации особенно актуальна для методов машинного обучения, широко используемых в системной биологии для выявления паттернов в многомерных сетевых данных. Сложные модели с большим числом параметров способны хорошо описывать обучающие данные, запоминая даже случайные флуктуации, но плохо предсказывают новые данные, поскольку выученные паттерны не отражают подлинных закономерностей. В высокоразмерных данных, характерных для системной биологии, где число переменных может превышать число наблюдений, риск переобучения особенно велик. Внешняя валидация на независимых наборах данных необходима для оценки реальной предсказательной силы моделей, но такие наборы не всегда доступны, и даже при их наличии воспроизводимость результатов между исследованиями часто низкая. Биомаркерные сигнатуры и классификаторы, идентифицированные в одном исследовании, нередко не воспроизводятся в других, что ставит под вопрос их биологическую обоснованность и клиническую применимость.
Проблема интерпретируемости результатов сетевого анализа создаёт барьер между статистическими паттернами и биологическим пониманием. Алгоритмы машинного обучения, особенно глубокие нейронные сети, могут выявлять сложные паттерны в данных и делать точные предсказания, но внутренняя логика этих предсказаний остаётся непрозрачной — это модели «чёрного ящика». Даже если модель точно предсказывает клинический исход, она не объясняет, почему этот исход происходит и какие механизмы его определяют. Для науки, стремящейся к пониманию, а не только к предсказанию, это серьёзное ограничение. Методы объяснимого искусственного интеллекта пытаются извлекать интерпретируемые правила из сложных моделей, но их успех ограничен. Кроме того, сетевые метрики, такие как централизация или модульность, являются абстрактными топологическими характеристиками, биологический смысл которых не всегда ясен: что означает для клетки иметь определённую центральность белка в интерактоме? Перевод сетевых описаний в механистическое понимание остаётся нерешённой задачей.
Редукция к структуре при игнорировании динамики представляет концептуальное ограничение многих сетевых подходов. Статическая карта сети — узлы и связи — не говорит, как сеть функционирует во времени, какие потоки информации и вещества через неё проходят, как она реагирует на возмущения. Две сети с идентичной топологией, но разными динамическими параметрами — скоростями реакций, порогами активации, константами связывания — могут демонстрировать качественно различное поведение. Анализ топологии необходим, но не достаточен для понимания сетевой функции; он должен дополняться динамическим моделированием, которое требует количественных данных о параметрах, часто недоступных или ненадёжных. Интеграция структурного и динамического анализа представляет собой важное направление развития системной биологии, но она сталкивается с вычислительными сложностями и дефицитом количественных данных.
Критика системной биологии включает также сомнения в её способности генерировать принципиально новое знание, а не только организовывать и визуализировать уже известное. Скептики указывают, что многие громкие утверждения системной биологии не привели к значимым открытиям или терапевтическим прорывам. Обещания персонализированной медицины, основанной на сетевом профилировании, остаются в значительной мере нереализованными спустя десятилетия после их провозглашения. Сетевые биомаркеры не заменили традиционных в клинической практике. Препараты, разработанные с использованием системно-биологических подходов, не демонстрируют принципиально лучшей эффективности, чем разработанные традиционно. Эта критика не означает, что системная биология бесполезна, но она указывает на разрыв между риторикой и реальными достижениями и призывает к более скромным и обоснованным ожиданиям.
Интеграция системного и редукционистского подходов представляется наиболее продуктивной стратегией, преодолевающей ограничения каждого подхода по отдельности. Редукционистские эксперименты генерируют детальное знание о свойствах отдельных компонентов, которое необходимо для параметризации и валидации сетевых моделей. Системные подходы интегрируют это знание и генерируют гипотезы о сетевых свойствах, которые затем тестируются редукционистскими экспериментами. Этот итеративный цикл между системным и редукционистским уровнями характеризует зрелую науку, которая использует сильные стороны обоих подходов. В исследованиях стресса это означает, что детальное изучение отдельных компонентов стрессового ответа — гормонов, рецепторов, сигнальных путей — остаётся необходимым и дополняется системным анализом их взаимодействий. Ни один подход в изоляции не даёт полного понимания; интеграция обоих — путь к прогрессу.
Связь обсуждения ограничений с содержанием курса состоит в формировании критического отношения к системно-биологическим концепциям и методам, представляемым в различных модулях. Понимание ограничений не отменяет ценности сетевых перспектив, но предохраняет от их абсолютизации и некритического принятия. Нулевой модуль о научной грамотности закладывает установку на критическую оценку всех научных утверждений, включая претензии системной биологии. Модуль о методах измерения обсуждает технические ограничения омиксных и нейровизуализационных методов, генерирующих сетевые данные. Модуль о передовых исследованиях рассматривает системную биологию в контексте её потенциала и ограничений. Понимание того, что системная биология — развивающееся поле с нерешёнными проблемами, а не завершённая методология с готовыми ответами, формирует адекватные ожидания и готовность к критическому взаимодействию с литературой.
Завершая обсуждение системной биологии как темы данного урока, необходимо подчеркнуть её статус перспективного, но незавершённого проекта в науке о стрессе. Парадигмальный сдвиг от молекул к сетям обоснован и продуктивен концептуально, но его методологическая реализация сталкивается с трудностями, которые ещё предстоит преодолеть. Сетевой подход предоставляет ценный концептуальный каркас для интеграции знаний о стрессе на разных уровнях и для понимания системной природы адаптации и патологии. Практические приложения сетевой медицины находятся на ранней стадии развития и требуют дальнейших разработок для клинического внедрения. Критическое осознание ограничений необходимо для продвижения вперёд и для предотвращения разочарования от нереалистичных ожиданий. Наука о стрессе, интегрирующая системные и редукционистские подходы, молекулярные и сетевые перспективы, находится на пути к более полному и действенному пониманию своего предмета.
5. Биопсихосоциальная модель: история и современность
5.1 Модель Энгеля 1977 года: революция и контекст
Публикация статьи Джорджа Энгеля «Необходимость новой медицинской модели: вызов биомедицине» в журнале Science в 1977 году стала поворотным моментом в истории медицинской мысли и заложила концептуальный фундамент для понимания здоровья и болезни, который продолжает определять теоретические рамки современной медицины и особенно психиатрии. Энгель, профессор психиатрии и терапии Рочестерского университета, сформулировал систематическую критику господствовавшей биомедицинской модели и предложил альтернативу, которую назвал биопсихосоциальной моделью. Эта публикация не была изолированным академическим упражнением; она кристаллизовала широкое недовольство редукционистским подходом к медицине, накопившееся в профессиональном сообществе к тому времени, и предоставила концептуальный язык для его артикуляции. Влияние статьи оказалось несоразмерным её объёму: она стала одной из наиболее цитируемых публикаций в истории медицины и превратила биопсихосоциальную модель в ортодоксию медицинского образования, особенно в области психиатрии, первичной медицинской помощи и психосоматической медицины.
Критика биомедицинской модели, составившая первую часть аргументации Энгеля, была направлена против редукционистского понимания болезни как исключительно биологического феномена, локализуемого в теле пациента и объяснимого нарушением структуры или функции органов, тканей, клеток, молекул. Биомедицинская модель, сформировавшаяся в девятнадцатом веке на волне успехов микробиологии, патологической анатомии и клеточной теории, постулировала, что болезнь представляет собой отклонение от нормы биологических показателей, и задача медицины состоит в идентификации и коррекции этого отклонения. В этой парадигме психологические и социальные факторы рассматривались либо как вторичные следствия биологического нарушения, либо как не относящиеся к собственно медицинской компетенции. Пациент редуцировался до носителя патологии, субъективный опыт болезни игнорировался как не поддающийся объективизации, контекст жизни пациента исключался из диагностического и терапевтического рассмотрения. Энгель утверждал, что эта модель, при всех её успехах в понимании и лечении инфекционных и многих соматических заболеваний, принципиально недостаточна для понимания здоровья и болезни в их полноте.
Аргументация Энгеля против биомедицинской модели опиралась на несколько линий критики, каждая из которых обнажала её ограничения. Первая линия касалась проблемы вариабельности: одна и та же биологическая аномалия у разных людей может проявляться по-разному или не проявляться вовсе; инфицирование патогеном не всегда приводит к болезни; многие люди с выраженными патоанатомическими изменениями не испытывают симптомов. Эта вариабельность указывает на то, что биологическая патология является необходимым, но не достаточным условием болезни; другие факторы определяют, проявится ли патология клинически. Вторая линия критики касалась субъективного измерения болезни: переживание боли, страдания, нетрудоспособности не тождественно биологическому нарушению и не может быть полностью из него выведено. Пациенты обращаются к врачу не с объективными биологическими показателями, а с субъективными жалобами и переживаниями, которые требуют понимания и адресации. Третья линия касалась социального контекста: заболеваемость, течение болезней, исходы лечения систематически различаются в зависимости от социального класса, культуры, семейного положения и других социальных факторов, которые не могут быть объяснены чисто биологическими различиями.
Альтернативная модель, предложенная Энгелем, постулировала, что здоровье и болезнь определяются взаимодействием факторов на трёх уровнях: биологическом, психологическом и социальном. Биологический уровень включает генетику, физиологию, биохимию, анатомию и патологию — всё то, что составляет предмет традиционной биомедицины. Психологический уровень охватывает когнитивные процессы, эмоции, поведение, личностные характеристики, стратегии совладания, субъективный опыт болезни. Социальный уровень включает межличностные отношения, семью, культуру, социально-экономические условия, системы здравоохранения и социальной поддержки. Болезнь в этой модели возникает не из изолированного нарушения на одном уровне, а из дисбаланса или дисфункции на пересечении уровней. Соответственно, понимание болезни и её лечение требуют внимания ко всем трём уровням, а не только к биологическому. Врач должен видеть не просто больной орган, а целого человека в контексте его психологической жизни и социальных отношений.
Теоретическим фундаментом биопсихосоциальной модели послужила общая теория систем, разработанная биологом Людвигом фон Берталанфи в середине двадцатого века. Общая теория систем предлагала универсальный концептуальный аппарат для описания сложных организованных целостностей в разных областях науки: от физики и биологии до психологии и социологии. Ключевым понятием этой теории была идея иерархии систем: элементарные частицы образуют атомы, атомы — молекулы, молекулы — органеллы, органеллы — клетки, клетки — ткани, ткани — органы, органы — организм, а организмы образуют социальные системы разного уровня. Каждый уровень этой иерархии обладает своими закономерностями, не сводимыми к нижележащему уровню, и требует специфических методов изучения. Энгель применил эту схему к медицине, позиционируя биопсихосоциальную модель как системный подход, интегрирующий уровни от молекулы до общества в единую концептуальную рамку понимания здоровья и болезни.
Исторический контекст появления биопсихосоциальной модели определялся несколькими интеллектуальными и социальными тенденциями семидесятых годов двадцатого века, которые создали рецептивную среду для её принятия. Холистическое движение в медицине, восходящее к традициям интегративной и гуманистической медицины, критиковало технологизацию и дегуманизацию здравоохранения, призывая к восстановлению отношений между врачом и пациентом как целостной личностью. Критика медикализации социальных проблем указывала на расширяющуюся тенденцию переопределять жизненные трудности, социальные отклонения и моральные вопросы как медицинские проблемы, подлежащие биологическому лечению. Интерес к психосоматике привлекал внимание к роли психологических факторов в развитии телесных заболеваний. Антипсихиатрическое движение подвергало радикальной критике биологический редукционизм психиатрии и её социальные функции. Социальная эпидемиология накапливала данные о социальных детерминантах здоровья. Все эти течения конвергировали к неудовлетворённости биомедицинской моделью и поиску альтернатив, которую биопсихосоциальная модель артикулировала и систематизировала.
Революционность биопсихосоциальной модели состояла не столько в оригинальности её отдельных положений, сколько в их систематической интеграции и легитимации в мейнстриме медицинского образования и практики. Идеи о роли психологических и социальных факторов в болезни высказывались и ранее: психосоматическая медицина, социальная медицина, медицинская антропология развивали эти темы на протяжении десятилетий. Однако эти направления оставались маргинальными по отношению к доминирующей биомедицинской парадигме и часто воспринимались как «мягкая» наука в противовес «твёрдой» биомедицине. Энгель, будучи одновременно терапевтом и психиатром с безупречной академической репутацией, публикуя в престижном естественнонаучном журнале, легитимировал интегративный подход и придал ему статус конкурирующей научной парадигмы, а не просто гуманистического призыва. Само название «биопсихосоциальная модель» стало мощным риторическим инструментом, обозначающим позицию холизма и интеграции в противовес редукционизму.
Влияние биопсихосоциальной модели на медицинское образование и практику оказалось глубоким и продолжительным, особенно в определённых дисциплинах и регионах. Психиатрия приняла биопсихосоциальную модель как официальную концептуальную рамку, и она вошла в учебники, программы резидентуры, клинические руководства. Первичная медицинская помощь, семейная медицина, поведенческая медицина также интегрировали модель в свои теоретические основания. Медицинские школы включили биопсихосоциальный подход в программы обучения, подчёркивая необходимость видеть пациента в контексте и развивать навыки коммуникации и эмпатии наряду с биомедицинскими компетенциями. Термин «биопсихосоциальный» стал практически обязательным в грантовых заявках, статьях и клинических описаниях, сигнализируя приверженность холистическому подходу. Однако, как будет показано в последующих разделах, разрыв между декларируемой приверженностью модели и её реальной реализацией в практике оказался значительным.
Связь биопсихосоциальной модели с содержанием данного курса о стрессе фундаментальна, поскольку сама структура курса отражает биопсихосоциальную логику организации знания. Модули о биологических механизмах стресса — нервной, эндокринной, иммунной регуляции — соответствуют биологическому компоненту модели. Модуль о психологии стресса — когнитивной оценке, эмоциях, копинге — реализует психологический компонент. Модуль о контекстах и популяциях рассматривает социальные детерминанты стресса. Сквозные связи между модулями воплощают принцип интеграции уровней. Настоящий урок, посвящённый уровням анализа и философским основаниям многоуровневого исследования, закладывает концептуальную базу для понимания биопсихосоциальной модели и её современных развитий. Однако курс также учитывает критику модели, рассматриваемую в последующих разделах, и стремится к более механистическому и операционализируемому пониманию связей между уровнями, чем предлагала оригинальная формулировка Энгеля.
Оценка исторической роли биопсихосоциальной модели требует признания как её вклада, так и её ограничений, которые станут предметом детального анализа далее. Модель Энгеля совершила важную работу критики и преодоления биомедицинского редукционизма, расширила горизонт медицинского мышления, легитимировала внимание к психологическим и социальным измерениям здоровья, повлияла на образование и самопонимание медицинских профессий. Вместе с тем она оставила нерешёнными фундаментальные методологические вопросы: как именно интегрировать уровни, каковы механизмы их взаимодействия, как операционализировать биопсихосоциальный подход для конкретных исследовательских и клинических задач. Эти нерешённые вопросы открыли пространство для критики, которая развивалась параллельно с институционализацией модели и которую необходимо рассмотреть для формирования сбалансированной оценки.
Завершая рассмотрение исторического контекста и содержания биопсихосоциальной модели Энгеля, следует подчеркнуть, что её появление представляло собой не изолированное интеллектуальное событие, а выражение более широкого сдвига в понимании сложных систем и многоуровневых феноменов. Этот сдвиг затронул не только медицину, но и психологию, социологию, экологию, организационные науки. Общая теория систем, кибернетика, теория сложности, эмерджентизм — все эти интеллектуальные движения разделяли интуицию о несводимости целого к частям и о необходимости интегративных подходов. Биопсихосоциальная модель была медицинским выражением этого более широкого интеллектуального климата. Понимание этого контекста помогает увидеть модель не как произвольную конструкцию одного автора, а как закономерный этап развития медицинской мысли, преодолевающей ограничения редукционизма и ищущей адекватные концептуальные рамки для сложных многоуровневых феноменов, каким является стресс и его последствия для здоровья.
5.2 Критика неопределённости и эклектизма
При всём влиянии и широком признании биопсихосоциальная модель с самого начала подвергалась систематической критике, центральным пунктом которой была её концептуальная неопределённость и недостаточная операционализируемость для научного исследования и клинической практики. Критики указывали, что модель постулирует важность биологических, психологических и социальных факторов для здоровья и болезни, но не специфицирует, как эти факторы взаимодействуют, каковы механизмы их связи, какова их относительная значимость при различных состояниях, как практически учитывать все три уровня в диагностике и лечении. Призыв «учитывать все факторы» звучит интуитивно убедительно, но не транслируется в конкретные методологические предписания. Модель оказывается настолько широкой и гибкой, что может вместить практически любое содержание и оправдать практически любой подход, что ставит под вопрос её научную ценность как руководства для исследования и практики.
Наиболее радикальная формулировка этой критики принадлежит американскому психиатру Нассиру Гаэми, который охарактеризовал биопсихосоциальную модель как «не теорию и не модель, а слоган» — риторическую формулу, создающую видимость научной позиции без её реального содержания. Теория в научном смысле предполагает набор связанных утверждений, из которых выводятся проверяемые предсказания; модель предполагает формализованное представление системы, позволяющее анализировать её поведение. Биопсихосоциальная модель, по мнению Гаэми, не удовлетворяет ни одному из этих критериев: она не генерирует специфических предсказаний, которые можно было бы опровергнуть; она не предлагает формализма, позволяющего количественно анализировать отношения между уровнями; она сводится к утверждению, что «всё важно», которое тривиально истинно и потому неинформативно. Такая критика, при всей её резкости, указывает на реальную методологическую слабость: модель, которую невозможно опровергнуть, не является научной теорией в попперианском смысле фальсифицируемости.
Проблема неопределённости проявляется прежде всего в отсутствии спецификации механизмов взаимодействия между уровнями. Биопсихосоциальная модель утверждает, что биологические, психологические и социальные факторы взаимодействуют, но не объясняет, как происходит это взаимодействие. Каким образом социальный фактор, например низкий социально-экономический статус, транслируется в биологическое изменение, например хроническое воспаление? Через какие психологические промежуточные переменные? Через какие биологические каскады? Модель оставляет эти вопросы без ответа, делегируя их эмпирическим исследованиям, но не предоставляя рамки для организации и интерпретации этих исследований. Это контрастирует с конкретными биологическими моделями, которые специфицируют молекулярные пути и позволяют генерировать проверяемые гипотезы о точках вмешательства. Критики указывают, что призыв к холизму без механистической конкретизации остаётся благим пожеланием, не направляющим ни исследование, ни практику.
Отсутствие методологии интеграции уровней представляет собой второй аспект критики неопределённости. Даже если признать, что все три уровня важны, остаётся вопрос: как практически их интегрировать в едином анализе? Какими методами изучать взаимодействие биологических, психологических и социальных переменных? Как строить модели, включающие переменные разного типа и разного уровня измерения? Биопсихосоциальная модель не предлагает ответов на эти методологические вопросы, и исследователи, работающие в её рамках, вынуждены изобретать методы заново или заимствовать их из других традиций. Проблемы многоуровневого моделирования, обсуждавшиеся во второй теме данного урока — операционализация конструктов, иерархический анализ, каузальная инференция — возникают именно здесь, но оригинальная формулировка модели не адресует их. Это создаёт разрыв между риторикой интеграции и реальностью дисциплинарно разобщённых исследований.
Проблема определения относительной значимости факторов также остаётся нерешённой в рамках биопсихосоциальной модели. Утверждение, что все три уровня важны, не говорит, какой из них важнее в конкретной ситуации, при конкретном расстройстве, для конкретного пациента. При одних состояниях биологические факторы могут доминировать настолько, что психологические и социальные интервенции оказываются малоэффективны без биологической коррекции. При других состояниях психосоциальные факторы могут быть настолько значимы, что биологическое вмешательство недостаточно или нерелевантно. Как определить, какой случай перед нами? Модель не предоставляет критериев для такого определения, оставляя это на клиническое суждение, которое может быть предвзятым, непоследовательным или определяться не научными соображениями, а дисциплинарной идентичностью клинициста. Психиатр с биологической ориентацией увидит биологические факторы; психолог — психологические; социальный работник — социальные. Модель, призванная интегрировать эти перспективы, не предоставляет оснований для такой интеграции.
Риск скатывания в эклектизм представляет собой практическое следствие концептуальной неопределённости модели. Эклектизм в данном контексте означает несистематическое комбинирование элементов из разных подходов без связующей теоретической рамки. Клиницист, декларирующий приверженность биопсихосоциальной модели, может на практике применять какой-то набор биологических, психологических и социальных интервенций, но не на основе интегрированного понимания их взаимодействия, а просто как список отдельных компонентов. Биологический психиатр назначает препарат и добавляет «рекомендацию поработать с психологом»; психотерапевт проводит терапию и отмечает «возможное влияние нейробиологии»; семейный врач спрашивает о социальной ситуации, но не знает, что делать с полученной информацией. Это формальное соблюдение буквы биопсихосоциального подхода при нарушении его духа: компоненты не интегрированы, а механически сложены. Реальная практика часто остаётся преимущественно биологической или преимущественно психологической, прикрытой риторикой холизма.
Использование биопсихосоциальной модели как риторического прикрытия представляет особенно проблематичный аспект её институционализации. Когда модель становится ортодоксией, декларируемой всеми, но реализуемой немногими, она превращается в ритуальную формулу, произносимую для демонстрации правильной позиции, но не определяющую реальную практику. Исследователь включает упоминание биопсихосоциальных факторов во введение статьи, но затем изучает только биологические или только психологические переменные. Клиницист проходит обучение, подчёркивающее целостность пациента, но в условиях временного давления и институциональных ограничений практикует так же, как практиковал бы без этого обучения. Медицинская школа декларирует биопсихосоциальную философию в своей миссии, но учебный план остаётся доминируемым биомедицинскими дисциплинами. Разрыв между риторикой и реальностью создаёт цинизм: модель воспринимается как благие пожелания, которые невозможно реализовать, и это восприятие становится самоисполняющимся пророчеством.
Конкретный пример эклектизма и неинтегрированности в практике можно наблюдать в лечении депрессии, которое декларируется биопсихосоциальным, но часто остаётся дуалистичным или даже монистичным. Биологический подход назначает антидепрессант, возможно, с кратким психообразованием, и считает свою задачу выполненной. Психотерапевтический подход проводит когнитивно-поведенческую или иную терапию, возможно, с рекомендацией консультации психиатра, но терапия не модифицируется на основе биологической информации. Комбинированное лечение сочетает препарат и терапию, но часто механически, без понимания того, как они взаимодействуют и потенцируют друг друга. Социальные интервенции — решение жилищных проблем, профессиональная реабилитация, работа с семьёй — рассматриваются как дополнительные к «настоящему» лечению, а не как его интегральная часть. Истинно интегрированный подход, который учитывал бы взаимодействие биологических, психологических и социальных факторов в формулировке случая и планировании лечения, остаётся скорее идеалом, чем реальностью.
Защитники биопсихосоциальной модели отвечают на критику неопределённости несколькими аргументами, заслуживающими рассмотрения. Первый аргумент состоит в том, что модель предназначена как метатеоретическая рамка, задающая общую ориентацию, а не как конкретная теория с детализированными предсказаниями; она не должна и не может заменить специфические теории на каждом уровне. Второй аргумент указывает, что неопределённость была неизбежна в 1977 году, когда знания о механизмах связи уровней были значительно менее развиты, и что современные исследования постепенно заполняют эти пробелы. Третий аргумент утверждает, что модель имела важную историческую функцию противодействия биологическому редукционизму, даже если её собственные позитивные предложения оставались расплывчатыми. Эти аргументы имеют определённую силу, но они не устраняют проблему: если модель остаётся неопределённой спустя почти полвека после её формулировки, это указывает на системную трудность, а не только на историческую незрелость.
Связь критики неопределённости с содержанием курса проявляется в стремлении курса преодолеть эту неопределённость через конкретизацию механизмов и методов. Модули о биологических механизмах стресса не просто констатируют важность биологии, а детально рассматривают молекулярные, клеточные и системные пути, через которые стресс влияет на организм. Модуль о психологии стресса специфицирует когнитивные и эмоциональные процессы, опосредующие стрессовую реакцию. Модуль о методах измерения предоставляет инструменты для операционализации переменных на каждом уровне. Связи между модулями указывают на конкретные механизмы взаимодействия уровней. Всё это представляет попытку наполнить биопсихосоциальную рамку конкретным содержанием, которого ей недоставало в оригинальной формулировке. Современные модели, рассматриваемые в последующих разделах данной темы, продолжают эту работу конкретизации.
Завершая обсуждение критики неопределённости и эклектизма, следует признать, что эта критика указывает на реальные методологические проблемы, которые не решаются простым призывом к холизму. Интеграция биологических, психологических и социальных факторов требует не только признания их важности, но и разработки конкретных методов их совместного изучения, моделирования их взаимодействия, операционализации для клинической практики. Эта работа ведётся, и прогресс достигается, но он не происходит автоматически из декларации биопсихосоциального подхода. Критическое осознание ограничений модели необходимо для её продуктивного развития и предохраняет от подмены реальной интеграции её риторической имитацией.
5.3 Критика игнорирования каузальных механизмов
Второй крупный кластер критики биопсихосоциальной модели касается её неспособности специфицировать каузальные отношения между факторами на разных уровнях, что превращает её скорее в описательную рамку корреляций, чем в объяснительную теорию причинно-следственных связей. Модель постулирует, что биологические, психологические и социальные факторы совместно определяют здоровье и болезнь, но не объясняет, как именно один фактор причинно влияет на другой, через какие механизмы и промежуточные переменные, в каком направлении и с какой силой. Утверждение, что низкий социально-экономический статус связан с повышенным уровнем стресса и худшими показателями здоровья, является эмпирически обоснованным, но оно описывает корреляцию, а не причинность. Для понимания и интервенции необходимо знать каузальные пути: через какие механизмы социальное неравенство транслируется в биологические изменения и болезнь. Биопсихосоциальная модель в её оригинальной формулировке не предоставляет таких каузальных путей, оставляя их спецификацию эмпирическим исследованиям без теоретического руководства.
Проблема каузальной неопределённости становится особенно острой при рассмотрении конкретных феноменов, которые биопсихосоциальная модель призвана объяснять. Рассмотрим связь социально-экономического статуса со здоровьем — один из наиболее устойчивых и хорошо документированных феноменов социальной эпидемиологии. Множество исследований демонстрируют градиент здоровья вдоль социальной иерархии: на каждой ступеньке социально-экономической лестницы здоровье в среднем лучше, чем на предыдущей. Биопсихосоциальная модель может описать этот феномен как результат взаимодействия социальных, психологических и биологических факторов, но это описание остаётся поверхностным без спецификации механизмов. Через какие пути низкий статус вредит здоровью? Финансовая нестабильность порождает хронический стресс, который через психологические процессы тревоги и руминации и биологические каскады кортизола и воспаления повреждает системы организма? Или низкий статус ограничивает доступ к качественному питанию, что напрямую влияет на метаболизм? Или низкий статус означает проживание в неблагоприятных условиях с загрязнением и шумом? Или низкий статус связан с дискриминацией и стигмой, которые через психологическую травму и хронический социальный стресс влияют на здоровье? Все эти пути вероятны, и все они могут действовать одновременно, но биопсихосоциальная модель не помогает различить их вклады и приоритизировать точки вмешательства.
Эпистемологическое значение каузальной спецификации состоит в том, что только каузальное знание позволяет эффективно вмешиваться и изменять исходы. Корреляционное знание сообщает, что переменные связаны, но не говорит, изменится ли одна при манипуляции другой. Если низкий социально-экономический статус коррелирует с плохим здоровьем, но не является его причиной, а обе переменные вызваны третьим фактором, то повышение статуса не улучшит здоровье. Если статус причинно влияет на здоровье, но через определённые пути, то только интервенция на эти пути будет эффективна. Без каузального понимания интервенции остаются слепыми попытками. Биопсихосоциальная модель, не специфицируя каузальные пути, не предоставляет руководства для разработки эффективных интервенций. Она указывает, что нужно учитывать социальные факторы, но не говорит, как именно вмешиваться в социальные детерминанты здоровья и какие из них приоритетны.
Проблема направленности каузальных связей между уровнями представляет дополнительную сложность, которую биопсихосоциальная модель не адресует. Являются ли причинные связи однонаправленными или двунаправленными? Влияет ли только социальное на биологическое, или биологическое также влияет на социальное? Очевидно, что связи двунаправленны: социальный стресс вызывает биологические изменения, но и биологические расстройства, например депрессия, приводят к социальным последствиям — потере работы, разрыву отношений, социальной изоляции. Существуют петли обратной связи, в которых каждый уровень влияет на другие и сам находится под их влиянием. Эти петли могут быть самоусиливающимися, создавая порочные круги, или самокорректирующимися, восстанавливая баланс. Модель, не специфицирующая направленность связей и характер обратных связей, не может предсказать динамику системы и точки наиболее эффективного вмешательства. Прерывание порочного круга может быть возможно в разных точках, но некоторые точки могут быть более доступны или эффективны, чем другие, и только каузальный анализ позволяет их идентифицировать.
Проблема опосредования и модерации каузальных связей добавляет ещё один слой сложности, требующей спецификации. Опосредование означает, что один фактор влияет на другой через промежуточную переменную: социальный стресс влияет на здоровье через воспаление, и воспаление является медиатором связи. Модерация означает, что сила связи между двумя факторами зависит от третьего: связь между стрессом и депрессией может быть сильнее при отсутствии социальной поддержки. Идентификация медиаторов указывает механизмы и возможные мишени вмешательства; идентификация модераторов указывает группы риска и защитные факторы. Биопсихосоциальная модель постулирует взаимодействие факторов, но не предоставляет систематической рамки для анализа медиации и модерации, не различает эти типы связей, не направляет их эмпирическое изучение. Это оставляет исследователей без теоретического руководства при планировании и интерпретации исследований механизмов.
Требование каузальных диаграмм как условия научной строгости было сформулировано современными критиками и методологами каузального вывода. Направленные ациклические графы, обсуждавшиеся в разделе о методах каузальной инференции, предоставляют формальный язык для репрезентации каузальных предположений: узлы представляют переменные, направленные рёбра — каузальные связи между ними. Такая диаграмма делает каузальные предположения явными и позволяет выводить условия их проверки: какие переменные контролировать, какие не контролировать, какие связи идентифицируемы из данных. Требование к теории сформулировать каузальную диаграмму представляет дисциплинирующее упражнение, обнажающее неопределённость и неполноту предположений. Биопсихосоциальная модель не может быть представлена каузальной диаграммой, поскольку она не специфицирует связи достаточно конкретно. Это не обязательно фатальный недостаток для метатеоретической рамки, но это ограничивает её полезность как руководства для конкретных исследований.
Проблема нефальсифицируемости, вытекающая из каузальной неопределённости, имеет серьёзные эпистемологические импликации для статуса биопсихосоциальной модели как научной теории. Согласно критерию Поппера, теория является научной в той мере, в которой она генерирует предсказания, которые могут быть опровергнуты наблюдением. Если любое наблюдение может быть интерпретировано как совместимое с теорией, теория не говорит ничего содержательного о реальности. Биопсихосоциальная модель уязвима для критики нефальсифицируемости: какой бы исход ни наблюдался, его можно пост-хок объяснить определённой комбинацией биологических, психологических и социальных факторов. Если терапия сработала, это потому что учли все факторы; если не сработала, значит какой-то фактор упустили. Такая модель защищена от опровержения, но ценой потери предсказательной силы. Это не означает, что модель бесполезна — она имеет эвристическую и организующую ценность, — но это означает, что она не функционирует как строгая научная теория.
Современные требования к механистическим объяснениям в биомедицине и психиатрии повышают стандарты, которым биопсихосоциальная модель в её оригинальной форме не соответствует. Механистическое объяснение специфицирует компоненты системы, их организацию и взаимодействия, благодаря которым система производит интересующий феномен. Объяснение депрессии через «биопсихосоциальные факторы» не является механистическим; объяснение через конкретную последовательность событий — хронический стресс активирует гипоталамо-гипофизарно-надпочечниковую ось, хроническое повышение кортизола снижает чувствительность глюкокортикоидных рецепторов в гиппокампе, это нарушает регуляцию оси и снижает нейрогенез, что способствует когнитивным и эмоциональным симптомам депрессии — является механистическим. Такие механистические модели тестируемы, фальсифицируемы, указывают мишени вмешательства. Биопсихосоциальная модель не предоставляет таких механизмов, хотя она совместима с ними и может служить рамкой для их организации.
Связь критики каузальной неопределённости с содержанием курса проявляется в акценте курса на механизмах, а не только на факторах. Модули о биологических механизмах стресса не ограничиваются констатацией, что биология важна; они детально рассматривают каузальные каскады — как психологическое восприятие угрозы транслируется в нейронную активность миндалины, как это активирует гипоталамус, как гипоталамус запускает гормональный каскад, как гормоны влияют на иммунные клетки и так далее. Модуль о психологии стресса специфицирует когнитивные процессы оценки и их связь с эмоциональными и биологическими ответами. Модуль о социальных контекстах рассматривает механизмы, через которые социальные условия транслируются в биологические изменения: концепция социальной геномики, обсуждаемая далее, является примером такой механистической спецификации. Курс стремится преодолеть абстрактную биопсихосоциальную рамку и наполнить её конкретным механистическим содержанием, отвечающим современным стандартам научного объяснения.
Проблемы методов установления причинности, обсуждавшиеся в разделе о каузальной инференции, приобретают здесь практическое значение. Для спецификации каузальных путей между биологическими, психологическими и социальными переменными необходимы методы, выходящие за пределы простых корреляционных исследований: лонгитюдные дизайны, устанавливающие временной порядок; экспериментальные и квазиэкспериментальные дизайны, манипулирующие предполагаемыми причинами; менделевская рандомизация, использующая генетические инструменты; каузальное моделирование с направленными ациклическими графами. Все эти методы активно применяются в современных исследованиях стресса и здоровья, постепенно заполняя каузальные пробелы биопсихосоциальной модели. Однако это заполнение происходит не благодаря модели, которая не предоставляет методологического руководства, а параллельно с ней, через развитие независимых методологических традиций.
Завершая обсуждение критики каузальной неопределённости, следует подчеркнуть, что эта критика не опровергает биопсихосоциальную модель, но указывает на необходимость её механистического наполнения. Признание важности биологических, психологических и социальных факторов — необходимый первый шаг, но недостаточный для объяснения и интервенции. Следующий шаг — спецификация каузальных путей между факторами, идентификация медиаторов и модераторов, построение тестируемых механистических моделей. Эта работа ведётся в современных исследованиях стресса, и некоторые её результаты рассматриваются в последующих разделах данной темы и в других модулях курса. Биопсихосоциальная модель может служить организующей рамкой для этой работы, но сама по себе она не заменяет механистического понимания и не освобождает от необходимости его достижения.
5.4 Критика недооценки биологического уровня
Парадоксальным образом биопсихосоциальная модель, первоначально предложенная как коррекция биологического редукционизма и призванная расширить медицинское мышление за пределы чистой биомедицины, в некоторых контекстах использовалась для противоположной цели — систематической недооценки или даже отрицания биологических факторов в пользу психосоциальных объяснений. Эта критика особенно актуальна для психиатрии, где исторические маятниковые колебания между биологическими и психосоциальными парадигмами создали условия для идеологизации биопсихосоциального подхода. Критики, в частности американский психиатр Генри Насралла, указывают, что модель, декларируя баланс между уровнями, на практике часто становилась инструментом для минимизации биологического компонента, особенно в руках клиницистов и теоретиков с преимущественно психосоциальной ориентацией. Эта тенденция была особенно выражена в период влияния антипсихиатрического движения и сохраняется в некоторых современных направлениях, что создаёт риск неадекватного лечения пациентов с расстройствами, требующими прежде всего биологической коррекции.
Историческая динамика парадигм в психиатрии двадцатого века создала контекст, в котором биопсихосоциальная модель могла интерпретироваться различными образами в зависимости от дисциплинарной позиции интерпретатора. Первая половина века была периодом доминирования психоанализа в американской психиатрии, когда психогенные объяснения преобладали и даже тяжёлые психозы рассматривались через призму ранних отношений и бессознательных конфликтов. Психофармакологическая революция пятидесятых-шестидесятых годов, открывшая эффективность хлорпромазина при шизофрении и имипрамина при депрессии, качнула маятник в биологическую сторону, и последующие десятилетия были периодом «биологической психиатрии» с акцентом на нейрохимию, генетику, нейровизуализацию. Антипсихиатрическое движение шестидесятых-семидесятых годов представляло радикальную реакцию на биологический редукционизм, отрицавшую саму реальность психических болезней как биологических сущностей. Биопсихосоциальная модель Энгеля появилась в этом контексте как попытка примирения и баланса, но её интерпретация неизбежно преломлялась через линзу этих исторических конфликтов.
Использование биопсихосоциальной модели для минимизации биологии проявляется в нескольких характерных формах, которые заслуживают критического анализа. Первая форма — это отрицание или преуменьшение нейробиологической основы тяжёлых психических расстройств, таких как шизофрения или биполярное расстройство. В наиболее радикальном варианте эти расстройства переопределяются как реакции на социальное угнетение, семейную дисфункцию или культурное конструирование «безумия», а биологические изменения, обнаруживаемые при них, интерпретируются как следствия, а не причины. Вторая форма — это систематическое предпочтение психосоциальных интервенций биологическим даже при состояниях, где последние продемонстрировали превосходящую эффективность. Третья форма — это риторическое использование биопсихосоциального языка для придания легитимности преимущественно психосоциальному подходу: клиницист упоминает биологию, но фактически работает только с психологическими и социальными факторами. Все эти формы представляют искажение интегративной идеи модели и могут вести к неоптимальному лечению.
Проблема биологической реальности психических расстройств приобретает особую остроту в контексте обсуждаемой критики. Накопленные за десятилетия исследования демонстрируют, что тяжёлые психические расстройства сопровождаются объективными биологическими изменениями на генетическом, молекулярном, клеточном, нейроанатомическом уровнях. Шизофрения имеет наследуемость порядка восьмидесяти процентов, сопровождается редукцией объёма серого вещества, изменениями дофаминергической нейротрансмиссии, нарушениями связности мозговых сетей. Биполярное расстройство характеризуется высокой наследуемостью, специфическими нейровизуализационными изменениями, эффективностью препаратов с известными биологическими механизмами. Отрицание или минимизация этих биологических данных во имя психосоциальной парадигмы представляет собой научный регресс, игнорирование эмпирических доказательств в пользу идеологических предпочтений. Биопсихосоциальная модель, правильно понятая, не отрицает эти биологические реальности, а помещает их в более широкий контекст, однако её расплывчатость позволяет интерпретации, которые de facto минимизируют биологию.
Практические последствия недооценки биологии для пациентов могут быть серьёзными и даже катастрофическими. Пациент с биполярным расстройством, который не получает стабилизаторов настроения из-за предпочтения клиницистом психотерапевтических подходов, рискует маниакальными эпизодами с разрушительными последствиями для его жизни, отношений, финансов, а также суицидом в депрессивной фазе. Пациент с шизофренией, которому задерживают антипсихотическую терапию в пользу психосоциальных интервенций, теряет критическое время, поскольку длительность нелеченого психоза ассоциирована с худшим долгосрочным прогнозом. Пациент с тяжёлой депрессией, которому предлагают только психотерапию, когда показана комбинация с фармакотерапией, страдает дольше и подвергается большему риску суицида. Эти клинические реальности требуют, чтобы биопсихосоциальный подход не становился прикрытием для идеологически мотивированного отказа от биологических интервенций там, где они необходимы.
Принцип дифференциального вклада факторов представляет собой необходимое дополнение к биопсихосоциальной модели, предотвращающее её искажение в сторону уравнивания всех уровней. Утверждение, что биологические, психологические и социальные факторы важны, не означает, что они одинаково важны при всех состояниях и у всех пациентов. При некоторых расстройствах биологические факторы настолько доминируют, что психосоциальные интервенции без биологической коррекции имеют ограниченную эффективность: это справедливо для большинства психозов, для тяжёлых депрессий, для биполярного расстройства. При других состояниях психосоциальные факторы могут быть центральными, и биологическое вмешательство либо не нужно, либо играет вспомогательную роль: это относится ко многим реактивным состояниям, адаптационным расстройствам, проблемам в отношениях. Ещё при других состояниях различные факторы могут быть действительно равнозначны, и интегрированное вмешательство на нескольких уровнях оптимально. Задача клинициста — определить конфигурацию факторов у конкретного пациента, а не механически применять идеологию равенства уровней.
Критерии для определения относительного вклада факторов остаются недостаточно разработанными, что отчасти объясняет сохраняющиеся споры. Тяжесть симптоматики традиционно считалась индикатором большего биологического вклада: тяжёлая депрессия с меланхолическими чертами скорее требует фармакотерапии, чем лёгкая. Семейная отягощённость и ранний возраст начала указывают на большую роль генетики. Отсутствие идентифицируемых психосоциальных триггеров при выраженной симптоматике также свидетельствует о преобладании биологии. Напротив, ясная связь симптомов с психосоциальными событиями, позднее начало, отсутствие семейной истории могут указывать на большую роль средовых факторов. Однако эти критерии вероятностны и не абсолютны; они направляют клиническое суждение, но не заменяют его. Биомаркеры, которые могли бы объективно определить биотип расстройства, остаются в стадии разработки. В отсутствие надёжных критериев клиническая практика неизбежно зависит от обучения, опыта и, к сожалению, идеологических предпочтений клинициста.
Интеграция не означает уравнивания — этот принцип должен быть центральным для адекватного понимания биопсихосоциальной модели. Признание множественных уровней детерминации не требует отрицания того, что при определённых состояниях один уровень может быть более релевантен, чем другие. Врач, лечащий диабет, учитывает психологические и социальные факторы — приверженность лечению, стресс, доступ к ресурсам, — но не отказывается от инсулина в пользу психотерапии. Аналогично, психиатр, лечащий шизофрению, учитывает психосоциальную реабилитацию, но не отказывается от антипсихотиков. Биопсихосоциальный подход означает внимание ко всем уровням, но не равное распределение терапевтических усилий между ними независимо от природы расстройства. Понимание этого принципа предохраняет от обоих крайностей: как от биологического редукционизма, игнорирующего психосоциальные факторы, так и от психосоциального редукционизма, минимизирующего биологию.
Исторические уроки недооценки биологии в психиатрии служат предостережением против повторения ошибок. Эра психоаналитического доминирования в американской психиатрии, когда шизофрения объяснялась «шизофреногенной матерью» и лечилась исключительно психоаналитически, привела к десятилетиям неэффективного лечения и страданий пациентов и их семей. Открытие эффективности антипсихотиков продемонстрировало, что биологическое вмешательство может принести облегчение там, где психотерапия бессильна. Аналогичные уроки были извлечены для депрессии и тревожных расстройств. Это не означает, что психотерапия бесполезна — напротив, она эффективна при многих состояниях и дополняет фармакотерапию при других. Но это означает, что отрицание или минимизация биологических факторов может иметь трагические последствия для пациентов. Биопсихосоциальная модель должна интегрировать эти исторические уроки, а не служить новым обоснованием для возврата к психосоциальному редукционизму.
Современные тенденции в психиатрии демонстрируют попытки преодоления идеологических крайностей и достижения подлинной интеграции. Понятие «биопсихосоциальной формулировки случая» предполагает систематический анализ вклада факторов всех уровней у конкретного пациента с составлением индивидуализированного плана лечения. Практика совместного принятия решений вовлекает пациента в обсуждение доступных опций и их соотношения. Развитие персонализированной медицины обещает более объективные критерии для определения оптимальных интервенций. Однако эти положительные тенденции сосуществуют с сохраняющимися идеологическими противостояниями, и полный консенсус о балансе факторов остаётся недостигнутым. Критическое осознание риска недооценки биологии, как и риска её переоценки, необходимо для поддержания подлинного баланса.
Связь обсуждаемой критики с содержанием курса проявляется в том, как курс позиционирует биологические и психосоциальные механизмы стресса. Модули о биологии стресса не представляют биологию как единственный или даже главный уровень; они представляют её как один из существенных уровней со своими специфическими механизмами. Модуль о психологии и модуль о социальных контекстах получают сопоставимое внимание, отражая подлинно интегративный подход. Модуль об интервенциях рассматривает биологические, психологические и социальные вмешательства, обсуждая показания для каждого и возможности их комбинации. Курс избегает обеих крайностей: он не редуцирует стресс к биологии и не минимизирует биологию в пользу психосоциальных факторов. Понимание того, что разные аспекты стресса и его последствий могут требовать акцента на разных уровнях, формирует гибкое клиническое мышление.
Завершая обсуждение критики недооценки биологии, следует подчеркнуть необходимость бдительности против идеологизации биопсихосоциального подхода в любом направлении. Модель была предложена как альтернатива редукционизму, но она может сама стать редукционистской, если используется для минимизации любого из её компонентов. Подлинная интеграция требует готовности признавать примат биологии там, где это обосновано данными, и примат психосоциальных факторов там, где обосновано это. Клиническое суждение, информированное эмпирическими данными и свободное от идеологических предубеждений, остаётся инструментом определения баланса в каждом конкретном случае. Биопсихосоциальная модель предоставляет рамку для такого суждения, но не заменяет его и не должна использоваться для оправдания пренебрежения любым из уровней.
5.5 Современные попытки формализации: механистические модели
Осознание ограничений оригинальной формулировки биопсихосоциальной модели — её неопределённости, эклектизма, каузальной неспецифичности — стимулировало современные исследовательские усилия по наполнению этой рамки конкретным механистическим содержанием. Эти усилия не отвергают биопсихосоциальный подход, а развивают его, предлагая специфические теории и модели, которые объясняют, как именно факторы разных уровней взаимодействуют, через какие механизмы социальный опыт транслируется в биологические изменения, какие психологические процессы опосредуют эту трансляцию. Результатом стало появление нескольких влиятельных теоретических конструкций, которые можно рассматривать как конкретизацию и операционализацию биопсихосоциальной интуиции. Эти модели различаются по фокусу и акцентам, но объединены стремлением преодолеть расплывчатость оригинальной формулировки и предоставить тестируемые гипотезы о механизмах связи между уровнями.
Модель аллостатической нагрузки, разработанная нейроэндокринологом Брюсом Макьюэном и его коллегами, представляет собой, возможно, наиболее влиятельную современную конкретизацию биопсихосоциального подхода к стрессу и здоровью. Центральная идея модели состоит в том, что хронический стресс, независимо от его источника — социального, психологического или физического, — активирует адаптивные физиологические системы, и при продолжительной активации эти системы изнашиваются, накапливая «аллостатическую нагрузку». Термин «аллостаз» обозначает процесс достижения стабильности через изменение — адаптивную перенастройку физиологических параметров в ответ на требования среды. Аллостатическая нагрузка — это кумулятивная цена этой адаптации, биологический износ, возникающий из хронического или повторяющегося стресса. Модель специфицирует конкретные биологические системы, накапливающие нагрузку, — нейроэндокринную, иммунную, метаболическую, сердечно-сосудистую — и предлагает операционализацию через измеримые биомаркеры: кортизол, адреналин, воспалительные маркеры, показатели метаболизма, кровяное давление, окружность талии.
Аллостатическая модель интегрирует уровни биопсихосоциальной рамки через спецификацию каузальных путей от социальных условий к биологическим исходам. Социальные факторы — низкий социально-экономический статус, дискриминация, небезопасное соседство, нестабильная занятость — порождают хронические стрессоры. Психологические процессы — когнитивная оценка угрозы, руминация, тревожное ожидание — определяют, как индивид воспринимает и обрабатывает эти стрессоры. Биологические системы активируются в ответ на воспринятый стресс и при хронической активации накапливают износ. Конкретность этих путей позволяет генерировать тестируемые гипотезы: если низкий социально-экономический статус увеличивает аллостатическую нагрузку, мы должны наблюдать повышенные уровни биомаркеров нагрузки у людей с низким статусом, и многочисленные исследования подтверждают это предсказание. Модель также специфицирует модераторы: индивидуальные различия в стрессовой реактивности, копинг-ресурсы, социальная поддержка могут буферировать накопление нагрузки при одинаковых объективных условиях.
Теория консервации ресурсов, разработанная психологом Стеваном Хобфоллом, предлагает альтернативную, но комплементарную конкретизацию биопсихосоциальной модели с акцентом на ресурсах и их утрате как центральном механизме стресса. Согласно этой теории, люди стремятся приобретать, сохранять и защищать ресурсы — объекты, условия, личные характеристики и энергии, которые они ценят или которые служат средством достижения ценных целей. Стресс возникает при угрозе утраты ресурсов, при их фактической утрате или при неспособности приобрести ожидаемые ресурсы после вложения других ресурсов. Ключевой принцип теории — примат потерь: утрата ресурсов психологически и биологически более значима, чем эквивалентное приобретение, и потери имеют тенденцию к каскадированию, когда одна потеря ведёт к другим. Эта теория объединяет психологическое измерение — субъективную значимость ресурсов, оценку угрозы и потери — с социальным измерением — объективной доступностью ресурсов в социальном контексте — и биологическим измерением — физиологическими затратами на совладание с потерями.
Интеграция теории консервации ресурсов с биологическими механизмами стресса демонстрирует возможности конкретизации биопсихосоциальных связей. Угроза значимым ресурсам активирует системы детекции угрозы — миндалину и связанные с ней структуры, которые в свою очередь запускают нейроэндокринные каскады стрессового ответа. Хроническая угроза или повторяющиеся потери ведут к хронической активации этих систем и накоплению аллостатической нагрузки. Социальный контекст определяет базовый уровень доступных ресурсов и уязвимость к потерям: люди с меньшими ресурсами более уязвимы, поскольку каждая потеря относительно значимее, и у них меньше возможностей для восстановления. Таким образом, теория объясняет социальный градиент здоровья через механизм ресурсной уязвимости и её биологических последствий. Этот уровень конкретности позволяет разрабатывать интервенции: укрепление ресурсов, предотвращение каскадов потерь, буферирование эффектов потерь через социальную поддержку и копинг-ресурсы.
Социальная геномика, направление исследований, развиваемое Стивом Коулом и коллегами, предоставляет наиболее молекулярно конкретную спецификацию того, как социальный опыт «проникает под кожу» и изменяет биологию на уровне экспрессии генов. Исследования в этой области показали, что социальные условия — одиночество, низкий социальный статус, ранняя социальная депривация — ассоциируются со специфическими паттернами экспрессии генов в иммунных клетках. Этот паттерн, названный «консервативным транскрипционным ответом на неблагополучие», характеризуется усилением экспрессии провоспалительных генов и ослаблением экспрессии генов противовирусного ответа. Биологическая логика этого паттерна, предположительно, связана с эволюционной адаптацией: в условиях социальной угрозы повышался риск раневых инфекций, требующих воспалительного ответа, в то время как в условиях социальной связности распространение вирусных инфекций было вероятнее, требуя противовирусного ответа. Социальный мозг считывает социальный контекст и через нейроэндокринные пути программирует иммунную систему на соответствующий режим.
Механизмы социальной геномики конкретизируют каузальный путь от социального опыта к молекулярным изменениям с беспрецедентной детальностью. Социальная изоляция или низкий статус воспринимаются нервной системой и активируют симпатоадреналовый ответ. Норадреналин и адреналин, высвобождаемые симпатическими нервами, иннервирующими лимфоидные органы, связываются с бета-адренергическими рецепторами на иммунных клетках. Активация этих рецепторов запускает внутриклеточные сигнальные каскады, которые модулируют активность транскрипционных факторов, включая ядерный фактор каппа-В и рецепторы глюкокортикоидов. Эти транскрипционные факторы регулируют экспрессию генов, определяя провоспалительный или противовоспалительный фенотип клетки. Таким образом, социальный сигнал транслируется в молекулярный язык экспрессии генов через определённую последовательность посредников. Эта конкретность позволяет идентифицировать потенциальные точки интервенции на каждом шаге каскада и генерировать тестируемые гипотезы о модераторах.
Модель воплощённого неравенства, или гипотеза «выветривания», предложенная Арлин Джеронимус, специфицирует, как социальное неравенство и дискриминация накапливают биологические последствия на протяжении жизни и даже поколений. Термин «выветривание» метафорически описывает преждевременное биологическое старение, которое наблюдается у людей, подвергающихся хроническому социальному стрессу, особенно у представителей дискриминируемых меньшинств. Согласно этой модели, хронический стресс от дискриминации, экономической нестабильности, жизни в неблагоприятных условиях накапливает аллостатическую нагрузку, которая проявляется в преждевременном старении физиологических систем, укорочении теломер, эпигенетических изменениях. Биологические последствия неравенства не просто коррелируют с социальной позицией, а причинно вытекают из хронического стрессового воздействия, связанного с этой позицией. Модель объясняет парадоксы, такие как ухудшение здоровья у афроамериканок среднего возраста несмотря на улучшение социально-экономического статуса: накопленное «выветривание» от десятилетий дискриминации не компенсируется поздними улучшениями.
Интеграция различных механистических моделей демонстрирует их комплементарность и возможность построения всеобъемлющей картины биопсихосоциальных механизмов стресса. Аллостатическая модель предоставляет общую рамку для понимания накопления биологического износа. Теория консервации ресурсов конкретизирует психологические механизмы, через которые социальные условия становятся субъективным стрессом. Социальная геномика раскрывает молекулярные пути трансляции социального сигнала в биологические изменения. Гипотеза выветривания объясняет кумулятивные эффекты на протяжении жизни. Каждая модель освещает определённый аспект общей картины, и вместе они наполняют биопсихосоциальную рамку конкретным механистическим содержанием, которого недоставало оригинальной формулировке Энгеля. Важно подчеркнуть, что эти модели не просто постулируют взаимодействие уровней, а специфицируют конкретные пути и механизмы этого взаимодействия, что делает их тестируемыми и фальсифицируемыми.
Связь механистических моделей с содержанием курса проявляется в организации материала различных модулей, которые детально рассматривают механизмы, обозначенные этими моделями. Первый модуль об организме как адаптивной системе вводит концепцию аллостатической нагрузки как центральную для понимания последствий хронического стресса. Четвёртый модуль об иммунитете и воспалении детально рассматривает консервативный транскрипционный ответ на неблагополучие и нейроиммунные механизмы социальной геномики. Пятый модуль о психологии стресса включает обсуждение теории консервации ресурсов в контексте когнитивных моделей стресса. Восьмой модуль о синдромах хронического стресса рассматривает межпоколенческую передачу травмы и накопление неблагоприятного опыта, связанные с гипотезой выветривания. Таким образом, курс не просто декларирует биопсихосоциальный подход, а конкретизирует его через рассмотрение специфических механистических моделей.
Перспективы дальнейшей формализации биопсихосоциального подхода связаны с развитием вычислительных методов, позволяющих моделировать сложные многоуровневые системы. Агентное моделирование позволяет симулировать, как индивиды с определёнными психологическими характеристиками взаимодействуют в социальном контексте и как это влияет на популяционные паттерны здоровья. Системная биология предоставляет инструменты для моделирования молекулярных сетей, опосредующих стрессовый ответ. Динамические модели в терминах дифференциальных уравнений позволяют формализовать аллостатические процессы и предсказывать траектории накопления нагрузки. Машинное обучение открывает возможности для идентификации паттернов в многомерных данных, охватывающих переменные всех уровней. Все эти методологические развития обещают дальнейшую формализацию биопсихосоциального подхода, однако их полная реализация — дело будущего, и текущие модели, при всей их конкретности, всё ещё оставляют значительные пробелы.
Завершая обсуждение современных механистических моделей, следует подчеркнуть, что они представляют существенный прогресс по сравнению с оригинальной биопсихосоциальной формулировкой. Эти модели переводят интуиции о важности множественных уровней в тестируемые гипотезы о конкретных механизмах. Они предоставляют основу для разработки интервенций, указывая потенциальные точки воздействия на каждом уровне. Они генерируют предсказания, которые могут быть опровергнуты эмпирически, соответствуя критериям научной теории. Вместе с тем эти модели не полны и не окончательны; они представляют собой этапы развивающегося понимания, которое продолжает уточняться и расширяться. Биопсихосоциальная модель, наполненная этим механистическим содержанием, становится более мощным инструментом для понимания и модификации связей между социальным опытом, психологическими процессами и биологическим здоровьем.
5.6 Альтернативные интегративные модели: сетевая теория и исследовательские критерии доменов
Помимо попыток модернизации и конкретизации биопсихосоциальной модели, в современной науке о психическом здоровье развиваются альтернативные интегративные подходы, которые предлагают иные концептуальные рамки для понимания психических расстройств и их связи с биологическими, психологическими и социальными факторами. Эти альтернативы не обязательно противоречат биопсихосоциальной модели, но предлагают отличные от неё способы концептуализации и операционализации многоуровневых связей. Две наиболее влиятельные альтернативы — сетевая теория психических расстройств и исследовательские критерии доменов — представляют различные, но равно амбициозные попытки преодолеть ограничения как традиционной биомедицинской модели, так и расплывчатой биопсихосоциальной рамки. Рассмотрение этих альтернатив расширяет понимание возможных подходов к интеграции уровней анализа и обогащает концептуальный репертуар для понимания стресса и его последствий.
Сетевая теория психических расстройств, разработанная нидерландским психологом Денни Борсбомом и коллегами, предлагает радикальное переосмысление природы психопатологии, которое отказывается от представления о симптомах как проявлениях скрытого заболевания в пользу понимания симптомов как узлов в причинной сети, активирующих друг друга. В традиционной медицинской модели симптомы являются следствиями болезни: жар, кашель и слабость — проявления инфекции, и лечение инфекции устраняет все симптомы. Сетевой подход предлагает альтернативу: симптомы психических расстройств могут причинно влиять друг на друга напрямую, без опосредования скрытой болезнью-сущностью. Бессонница вызывает усталость, усталость снижает активность, снижение активности ведёт к социальной изоляции, изоляция усиливает руминацию, руминация углубляет подавленное настроение, и так далее. Каждая стрелка в этой сети представляет собой причинный механизм, который может быть биологическим, психологическим или социальным. Расстройство в этой модели — не сущность, вызывающая симптомы, а самоподдерживающийся паттерн активации в сети симптомов.
Сетевая теория интегрирует биологические, психологические и социальные механизмы иным способом, чем биопсихосоциальная модель, — не через постулирование уровней, а через спецификацию связей между симптомами. Связь между бессонницей и усталостью имеет биологические механизмы: нарушение восстановительных процессов сна. Связь между социальной изоляцией и руминацией имеет психологические механизмы: отсутствие внешних стимулов, направляющих внимание вовне. Связь между руминацией и подавленным настроением имеет как психологические, так и биологические аспекты: негативное мышление и связанные с ним нейронные паттерны. Таким образом, биопсихосоциальное содержание не теряется, а рассредоточивается по связям сети. Каждая связь может быть мишенью интервенции, и разрыв критических связей может дестабилизировать патологический паттерн. Эта модель естественно объясняет коморбидность: если симптомы расстройств связаны друг с другом, одно расстройство может активировать симптомы другого через мостовые связи.
Преимущества сетевого подхода включают его операционализируемость и тестируемость, которых недоставало биопсихосоциальной модели. Сетевой анализ использует математический аппарат теории графов и позволяет эмпирически оценивать структуру сетей симптомов на основе данных о корреляциях и временных последовательностях. Можно идентифицировать центральные симптомы — узлы с наибольшим числом связей или с наибольшим влиянием на сеть, — которые являются потенциальными приоритетными мишенями интервенции. Можно оценивать связность сети — показатель, который может отражать уязвимость к развитию расстройства: более связная сеть легче активируется и труднее деактивируется. Можно моделировать распространение активации по сети и предсказывать, какие паттерны симптомов с большей вероятностью возникнут при определённых триггерах. Вся эта методология количественна, воспроизводима и позволяет генерировать фальсифицируемые предсказания, что отвечает стандартам научной теории.
Критика сетевого подхода указывает на ряд ограничений, которые необходимо учитывать при оценке его потенциала как альтернативы или дополнения к биопсихосоциальной модели. Первое ограничение — риск атеоретичности: сетевой анализ может идентифицировать корреляционные структуры в данных без понимания биологических или психологических механизмов, лежащих за этими корреляциями. Статистически идентифицированная связь между симптомами не эквивалентна пониманию того, почему и как один симптом вызывает другой. Второе ограничение — техническая чувствительность результатов к выбору симптомов, включаемых в анализ, к методам оценки связей, к статистическим порогам. Третье ограничение — недостаточная клиническая валидация: хотя сетевые модели теоретически обоснованы, эмпирические демонстрации того, что интервенции, основанные на сетевом анализе, более эффективны, чем традиционные подходы, пока ограничены. Четвёртое ограничение — сетевой подход лучше описывает поддержание расстройств, чем их этиологию; он объясняет, как симптомы усиливают друг друга, но не всегда объясняет, что первично активировало сеть.
Исследовательские критерии доменов, известные под аббревиатурой RDoC от английского «Research Domain Criteria», представляют собой инициативу Национального института психического здоровья США, направленную на реорганизацию исследований психопатологии вокруг биологически обоснованных измерений вместо традиционных диагностических категорий. Отправной точкой инициативы послужила неудовлетворённость традиционными диагностическими системами, такими как DSM, которые основаны на синдромальных описаниях и не обязательно отражают биологическую реальность. Диагноз «большое депрессивное расстройство» может объединять пациентов с совершенно различными биологическими механизмами, что затрудняет исследование и персонализацию лечения. RDoC предлагает альтернативу: вместо исследования болезней как унитарных сущностей исследовать трансдиагностические домены функционирования — такие как негативная валентность, позитивная валентность, когнитивные системы, социальные процессы — на всех уровнях анализа, от генов до поведения.
Структура RDoC организована как матрица, в которой строки представляют домены и конструкты, а столбцы — уровни анализа. Домены включают системы негативной валентности (страх, тревога, потеря), системы позитивной валентности (вознаграждение, мотивация), когнитивные системы (внимание, память, когнитивный контроль), системы социальных процессов (привязанность, коммуникация, понимание себя и других), системы возбуждения и регуляции (циркадные ритмы, сон, возбуждение). Каждый домен подразделяется на более специфические конструкты. Уровни анализа охватывают диапазон от молекул и генов через клетки, цепи, физиологию, поведение до самоотчёта. Эта матрица задаёт систематическую рамку для многоуровневых исследований: каждый конструкт изучается на всех уровнях, и связи между уровнями становятся эксплицитным предметом исследования. В отличие от расплывчатого призыва биопсихосоциальной модели «учитывать все факторы», RDoC предлагает конкретную структуру для организации многоуровневого исследования.
Преимущества подхода RDoC для интеграции уровней анализа включают его систематичность, операционализируемость и ориентацию на биологию. Систематичность проявляется в явной организации доменов и уровней, которая направляет исследовательские усилия и обеспечивает сопоставимость результатов разных исследований. Операционализируемость обеспечивается тем, что каждый конструкт определяется через конкретные измерения на каждом уровне: конструкт страха операционализируется через определённые гены, нейронные цепи, физиологические реакции, поведенческие паттерны, субъективные переживания. Ориентация на биологию отражает стремление укоренить психологические конструкты в нейробиологии, что обещает более тесную связь с разработкой биологических методов лечения. Для исследований стресса RDoC предоставляет рамку для изучения того, как стресс влияет на различные домены функционирования — усиливает негативную валентность, ослабляет позитивную, нарушает когнитивный контроль — и как эти эффекты реализуются на разных уровнях.
Критика подхода RDoC включает несколько существенных возражений, которые ограничивают его применимость как полной замены традиционных и биопсихосоциальных подходов. Первое возражение касается редукционизма: критики утверждают, что RDoC возвращает нас к биологическому редукционизму, от которого биопсихосоциальная модель стремилась уйти, поскольку он ориентирован на поиск биологических оснований психологических конструктов и недостаточно учитывает социальные детерминанты. Социальный уровень анализа в матрице RDoC отсутствует или представлен минимально. Второе возражение связано с клинической применимостью: RDoC разработан как исследовательская рамка, а не как клинический инструмент, и неясно, как транслировать его конструкты в диагностику и лечение конкретных пациентов. Третье возражение касается зрелости рамки: несмотря на десятилетие развития, RDoC не привёл к ожидавшимся прорывам в понимании и лечении психических расстройств, и его практическая ценность остаётся предметом дискуссии.
Сравнение альтернативных подходов с биопсихосоциальной моделью выявляет их взаимодополняющие сильные стороны и ограничения. Биопсихосоциальная модель предлагает широкую философскую ориентацию на интеграцию уровней, но не предоставляет конкретной методологии. Сетевой подход предлагает операционализируемую методологию, но сфокусирован на симптоматическом уровне и может упускать этиологию. RDoC предлагает систематическую многоуровневую структуру, но смещён в сторону биологии и недостаточно клинически ориентирован. Ни один из этих подходов не является полным и самодостаточным; каждый освещает определённые аспекты сложной реальности психических расстройств и их связи с биологическими, психологическими и социальными факторами. Продуктивная стратегия может состоять не в выборе одного подхода, а в их интеграции: биопсихосоциальная рамка задаёт общую ориентацию, сетевой подход моделирует взаимодействие симптомов, RDoC организует изучение базовых механизмов.
Перспективы развития интегративных моделей связаны с преодолением ограничений каждого из существующих подходов и их конвергенцией. Биопсихосоциальная модель нуждается в дальнейшей механистической конкретизации, которую предоставляют модели, обсуждавшиеся в предыдущем разделе. Сетевой подход нуждается в углублении теоретического понимания механизмов связей и в клинической валидации. RDoC нуждается в расширении социального измерения и в разработке путей к клиническому применению. Появляющиеся синтетические подходы пытаются объединить сильные стороны различных рамок: сетевые модели, включающие биологические, психологические и социальные узлы; многоуровневые модели в духе RDoC, учитывающие социальный контекст; механистические спецификации биопсихосоциальных связей с использованием вычислительных методов. Поиск оптимальной интегративной модели продолжается, и текущий момент характеризуется продуктивным плюрализмом подходов.
Связь альтернативных моделей с содержанием курса проявляется в нескольких модулях. Сетевой подход к психопатологии обсуждается в модуле о психических расстройствах, связанных со стрессом, в контексте понимания коморбидности и трансдиагностических процессов. Многоуровневая структура RDoC созвучна организации курса, который рассматривает стресс на уровнях от молекулярного до социального. Модуль о передовых исследованиях включает обсуждение сетевой науки и вычислительной психиатрии как перспективных направлений. Домены RDoC — негативная валентность, позитивная валентность, когнитивные системы — соответствуют темам, рассматриваемым в модуле о психологии стресса. Таким образом, альтернативные интегративные модели не противопоставляются содержанию курса, а обогащают его концептуальный арсенал.
Завершая обсуждение альтернативных интегративных моделей и темы биопсихосоциального подхода в целом, следует подчеркнуть, что поиск адекватных концептуальных рамок для понимания стресса и его последствий продолжается и представляет собой одну из центральных задач современной науки о здоровье и болезни. Биопсихосоциальная модель Энгеля сыграла историческую роль в преодолении биологического редукционизма и легитимации многоуровневого подхода. Критика модели выявила её ограничения: неопределённость, каузальную неспецифичность, риск эклектизма и идеологизации. Современные механистические модели — аллостатическая нагрузка, консервация ресурсов, социальная геномика — наполняют рамку конкретным содержанием. Альтернативные подходы — сетевая теория, RDoC — предлагают иные способы организации многоуровневого понимания. Интеграция этих развитий в когерентную и операционализируемую теорию стресса и здоровья остаётся задачей для будущих исследований, и данный курс стремится вносить вклад в эту задачу через систематическое изложение механизмов на всех уровнях анализа.
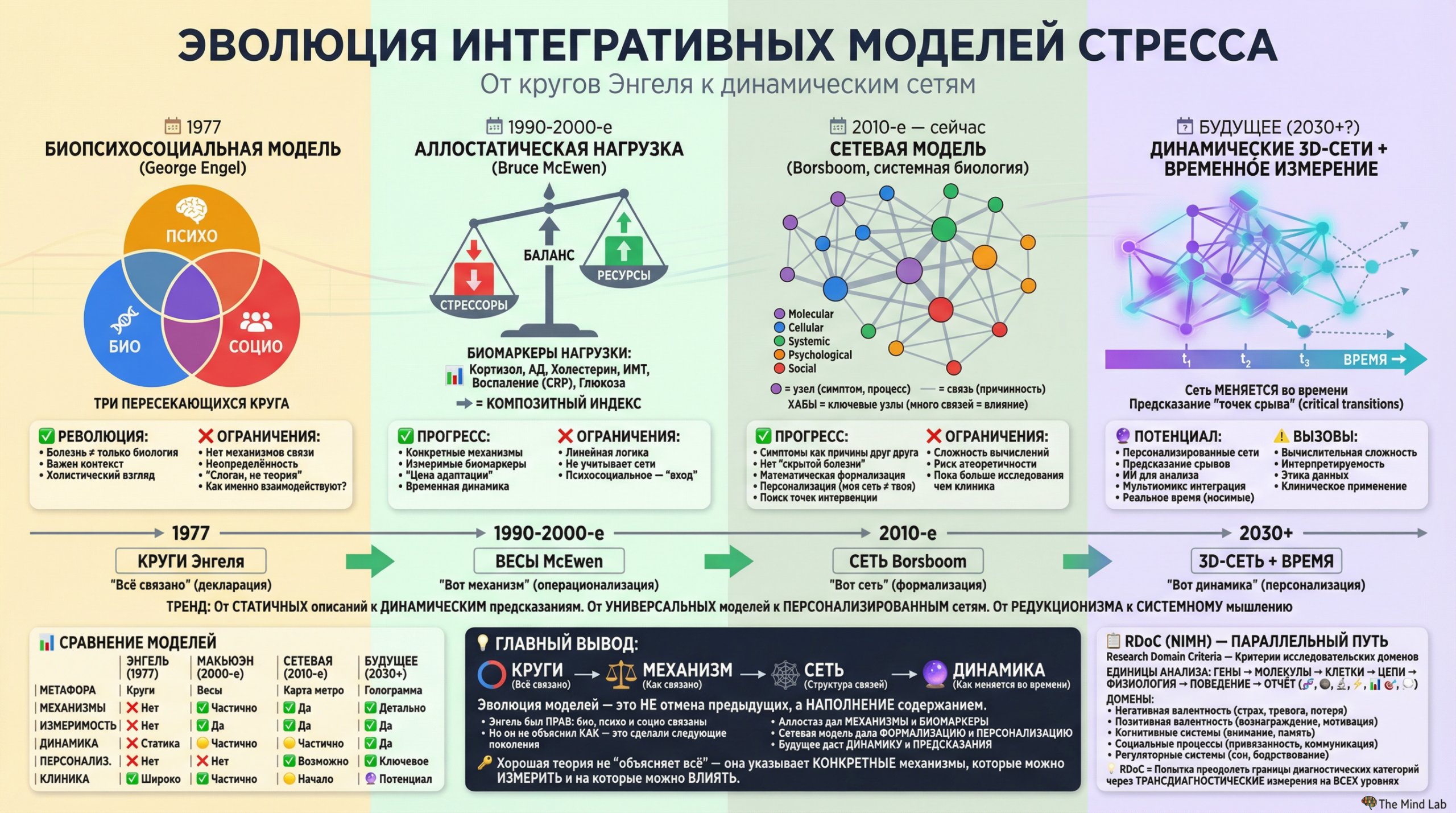
Интегрирующее обсуждение: философия и практика многоуровневого исследования
Эпистемологический плюрализм как необходимость
Завершающее рассмотрение философских и методологических оснований науки о стрессе приводит к фундаментальному выводу о необходимости эпистемологического плюрализма как единственной адекватной позиции для понимания и исследования этого сложного многоуровневого феномена. Эпистемологический плюрализм в данном контексте означает признание того, что полноценное понимание стресса требует множественных перспектив, методов, уровней анализа и теоретических рамок, которые не сводимы друг к другу, но взаимно дополняют и ограничивают друг друга. Это не релятивистская позиция, утверждающая равную ценность любых точек зрения; это обоснованный методологический выбор, вытекающий из самой природы изучаемого феномена. Стресс конституирован одновременно на молекулярном, клеточном, системном, психологическом и социальном уровнях, и каждый из этих уровней обладает своими закономерностями, несводимыми к другим уровням, но связанными с ними сложными отношениями детерминации. Ни один уровень анализа, ни одна дисциплина, ни один метод не способны схватить стресс во всей его полноте; только их интеграция может приблизить нас к адекватному пониманию.
Стресс как феномен обладает онтологическими характеристиками, которые делают монистические эпистемологические стратегии принципиально недостаточными независимо от уровня, на котором они применяются. Попытка понять стресс исключительно через молекулярную биологию упускает смыслы и контексты, определяющие субъективное переживание и социальную детерминацию стрессоров. Попытка понять стресс исключительно через психологию упускает биологические механизмы, реализующие психологические процессы и определяющие их ограничения. Попытка понять стресс исключительно через социологию упускает индивидуальную вариабельность биологических и психологических ответов на социальные условия. Каждый монистический подход схватывает реальный аспект стресса, но оставляет за пределами рассмотрения другие, столь же реальные аспекты. Это не вопрос предпочтений или дисциплинарных традиций; это следствие многоуровневой природы самого феномена, который не может быть адекватно репрезентирован ни в одной отдельной перспективе. Признание этой онтологической многоуровневости влечёт эпистемологический плюрализм как методологический императив.
Редукционизм, рассмотренный в первой теме данного урока, занимает необходимое, но ограниченное место в плюралистической эпистемологии науки о стрессе. Его необходимость обусловлена тем, что понимание механизмов на фундаментальных уровнях создаёт основу для объяснения и интервенции: знание того, как кортизол связывается с глюкокортикоидным рецептором, как рецептор регулирует транскрипцию генов, как изменение экспрессии генов влияет на функцию нейронов — всё это представляет собой механистическое знание, без которого понимание стресса осталось бы поверхностным и описательным. Редукционистская программа генерирует это знание с беспрецедентной точностью и детальностью, и её достижения составляют фундамент современной науки о стрессе. Однако ограниченность редукционизма столь же фундаментальна: знание механизмов на нижележащих уровнях не транслируется автоматически в понимание феноменов на вышележащих уровнях. Полное знание молекулярных каскадов не предсказывает субъективное переживание тревоги; полное знание нейронных цепей не объясняет социальные паттерны распространённости депрессии. Редукция к элементам теряет эмерджентные свойства целого, которые столь же реальны и столь же нуждаются в объяснении.
Эмерджентность, обсуждавшаяся как концептуальная альтернатива полному редукционизму, представляет собой не мистическое понятие и не отрицание научного объяснения, а признание того, что определённые свойства возникают из организации и взаимодействия компонентов и требуют изучения на своём уровне. Синхронизация активности нейронных ансамблей — эмерджентное свойство, не локализованное ни в одном нейроне. Руминация как когнитивный паттерн — эмерджентное свойство взаимодействия мозговых сетей, не сводимое к активности отдельных нейронов или молекул. Социальная поддержка как защитный фактор — эмерджентное свойство сети межличностных отношений. Признание эмерджентности не означает отказа от поиска объяснений; оно означает признание того, что объяснения эмерджентных феноменов должны включать понимание того, как они возникают из взаимодействий, но не могут быть полностью заменены описанием этих взаимодействий. Нужны законы и закономерности на уровне эмерджентных феноменов, которые дополняют, а не заменяют законы нижележащих уровней. Эпистемологический плюрализм включает множественность уровней объяснения как необходимое следствие онтологической эмерджентности.
Нисходящая причинность, рассмотренная как механизм влияния высших уровней на низшие, дополнительно обосновывает необходимость плюрализма, демонстрируя, что причинные связи в биологических системах не являются однонаправленными снизу вверх. Когнитивная переоценка угрозы, являющаяся психологическим процессом, изменяет активность миндалины и секрецию кортизола, которые являются биологическими процессами. Социальная поддержка, являющаяся социальным феноменом, модулирует иммунную функцию и экспрессию генов. Эти нисходящие влияния реальны, измеримы, воспроизводимы и имеют практическое значение для терапии. Они демонстрируют, что высшие уровни не являются лишь эпифеноменами низших, но обладают собственной каузальной силой. Эпистемология, игнорирующая нисходящую причинность, упускает существенный аспект реальности и не способна объяснить эффективность психологических и социальных интервенций. Плюралистическая эпистемология, признающая причинность на всех уровнях и в обоих направлениях, адекватнее отражает сложную структуру биопсихосоциальной детерминации.
Множественная реализуемость, обсуждавшаяся как философская концепция, имеющая прямые эмпирические проявления в гетерогенности стресс-связанных расстройств, представляет собой ещё один аргумент в пользу эпистемологического плюрализма. Один и тот же клинический синдром — депрессия, тревожное расстройство, посттравматическое стрессовое расстройство — может быть реализован через различные биологические механизмы у разных пациентов. Это означает, что психологический уровень описания, на котором формулируется диагноз, не редуцируется однозначно к биологическому уровню; отношение между уровнями сложнее, чем отношение однозначного соответствия. Одна и та же психология может иметь различную биологию, и понимание этого многообразия требует исследования на обоих уровнях. Монистическая эпистемология, сводящая психологическое к биологическому, упускает эту множественность и ведёт к ошибочным ожиданиям универсальных биологических маркеров и единственных механизмов для категориальных диагнозов. Плюралистическая эпистемология признаёт множественность реализаций и направляет исследования на выявление биотипов внутри диагностических категорий.
Многоуровневое моделирование, обсуждавшееся во второй теме урока, представляет собой методологическое воплощение эпистемологического плюрализма и одновременно демонстрирует его практические трудности. Идеал многоуровневого исследования состоит в одновременном измерении и анализе переменных на нескольких уровнях с изучением связей между ними. Однако реализация этого идеала сталкивается с проблемами: определение границ уровней концептуально неоднозначно, операционализация конструктов на разных уровнях порождает проблему концептуальной эквивалентности, статистические методы требуют больших выборок и сложных моделей, временная динамика плохо схватывается кросс-секционными дизайнами, интеграция разнородных омиксных данных вычислительно интенсивна, каузальная инференция в наблюдательных данных ограничена. Эти трудности не опровергают плюралистический идеал, но указывают на то, что его реализация требует методологических инноваций и не достигается простым декларированием многоуровневого подхода. Признание этих трудностей является частью эпистемологической скромности, которая должна сопровождать плюралистическую позицию.
Трансляционный разрыв, рассмотренный в третьей теме, представляет собой практическое свидетельство того, что монистический редукционизм доклинической науки недостаточен и что сложность человеческого стресс-ответа систематически недооценивается при экстраполяции из модельных систем. Высокий процент неудач при переносе препаратов из доклинических испытаний в клиническую практику отражает разрыв между редукционистским пониманием механизмов на животных моделях и интегративной сложностью человеческого организма с его генетической гетерогенностью, уникальной психологией, социальным контекстом. Трансляционный разрыв — это не случайность и не технический сбой, который можно устранить лучшими моделями; это индикатор фундаментального ограничения редукционистской стратегии, которая изолирует механизмы из их многоуровневого контекста. Плюралистический подход, учитывающий человеческую гетерогенность, психологические и социальные модераторы, индивидуальные траектории, обещает сократить этот разрыв, хотя и ценой большей сложности исследований.
Ограничения животных моделей, подробно обсуждавшиеся в контексте трансляционного разрыва, иллюстрируют межвидовые различия, которые делают невозможным простой перенос результатов с грызунов на людей. Эти ограничения не являются аргументом против использования животных моделей, которые остаются незаменимыми для определённых целей; они являются аргументом против монистической эпистемологии, которая считает животные данные достаточными для понимания человеческого стресса. Плюралистическая позиция включает животные модели как один из источников знания, дополняемый человеческими клеточными моделями, клиническими наблюдениями, эпидемиологическими данными, психологическими исследованиями. Триангуляция этих разнородных источников повышает надёжность выводов, поскольку каждый источник имеет свои ограничения, но они не совпадают, и конвергенция результатов из разных источников указывает на робастные закономерности.
Гетерогенность человеческих популяций, также обсуждавшаяся как источник трансляционных трудностей, представляет собой не помеху для науки, которую нужно устранить стандартизацией, а существенную характеристику реальности, которую нужно понять и учесть. Генетическое разнообразие, эпигенетические различия, вариации жизненного опыта, культурные контексты создают популяции, в которых одни и те же вмешательства дают различные результаты у разных индивидов. Монистическая эпистемология, ищущая универсальные законы, применимые ко всем, систематически недооценивает эту гетерогенность и терпит неудачу при столкновении с ней в клинических испытаниях. Плюралистическая эпистемология рассматривает гетерогенность как информацию о структуре феномена, ищет биотипы и подгруппы, разрабатывает персонализированные подходы. Это не отказ от обобщений, а их уточнение: законы формулируются не для абстрактного среднего человека, а для определённых конфигураций факторов.
Системная биология, рассмотренная в четвёртой теме, предлагает новый концептуальный и методологический аппарат для интеграции знаний о стрессе, который преодолевает некоторые ограничения традиционного редукционизма. Сетевой подход, рассматривающий организм как систему взаимодействующих компонентов, схватывает эмерджентные свойства, возникающие из организации связей, а не только из свойств элементов. Молекулярные сети, нейронные сети, динамические системы предоставляют формализмы для моделирования сложных взаимодействий на разных уровнях. Однако системная биология, при всём её потенциале, не является панацеей и не заменяет плюрализм монистической альтернативой. Сетевые модели сами по себе требуют данных из редукционистских экспериментов для параметризации; они не охватывают психологические и социальные уровни без их явного включения; они сталкиваются с собственными ограничениями вычислительной сложности, неполноты данных, интерпретируемости. Системная биология — мощный инструмент в плюралистическом арсенале, но не его замена.
Сетевая медицина, как приложение системной биологии к терапии, иллюстрирует, как плюралистическое понимание болезни может транслироваться в практику. Признание того, что болезнь представляет собой возмущение в сети взаимодействующих компонентов, а не поломку одного элемента, ведёт к стратегиям полифармакологии, комбинированной терапии, репозиционирования препаратов. Эти стратегии воплощают плюралистическую логику: воздействие на несколько точек сети одновременно, учёт сетевых эффектов и компенсаторных механизмов, персонализация на основе индивидуального сетевого профиля. Вместе с тем сетевая медицина остаётся преимущественно на биологическом уровне и требует дополнения психологическими и социальными интервенциями для полноценной реализации биопсихосоциального подхода. Плюрализм в терапии означает не только комбинацию биологических мишеней, но и комбинацию уровней вмешательства.
Теория сложных систем добавляет к сетевому пониманию динамическое измерение, вводя концепции аттракторов, критических переходов, устойчивости, которые трансформируют понимание здоровья и болезни. Здоровье и болезнь в этой перспективе — не точки на континууме, а качественно различные аттракторы, устойчивые состояния, между которыми возможны резкие переходы при постепенном изменении условий. Эта динамическая перспектива дополняет статическое сетевое описание и открывает новые возможности для предсказания и превенции срывов через идентификацию предвестников критических переходов. Однако теория сложных систем, как и системная биология, не исчерпывает плюралистическую программу; она предоставляет математический язык для описания динамики, но сама по себе не специфицирует биологические, психологические или социальные механизмы, реализующие эту динамику. Интеграция динамической перспективы с механистическим пониманием на каждом уровне представляет задачу для плюралистической науки о стрессе.
Биопсихосоциальная модель, детально проанализированная в пятой теме, представляет собой раннюю артикуляцию плюралистической интуиции, значение которой не умаляется критикой её ограничений. Энгель правильно увидел, что биологический редукционизм недостаточен и что здоровье и болезнь определяются взаимодействием факторов на нескольких уровнях. Эта интуиция остаётся справедливой и составляет концептуальное ядро плюралистической эпистемологии. Однако оригинальная формулировка модели страдала неопределённостью, не специфицируя механизмы связи уровней, методы их изучения, критерии определения относительной значимости. Эта неопределённость открывала возможности для эклектизма, риторического использования, идеологизации. Развитие плюралистического подхода требует преодоления этих ограничений через механистическую конкретизацию, которую предоставляют модели аллостатической нагрузки, консервации ресурсов, социальной геномики, рассмотренные в ходе урока.
Механистические модели, наполняющие биопсихосоциальную рамку конкретным содержанием, представляют прогресс в направлении операционализируемого плюрализма. Модель аллостатической нагрузки специфицирует, как хронические стрессоры любого происхождения — социального, психологического, физического — через психологическую оценку и биологические каскады накапливают измеримый биологический износ. Теория консервации ресурсов объясняет психологическую динамику стресса через угрозу и утрату ценных ресурсов с каскадирующими эффектами. Социальная геномика раскрывает молекулярные пути, через которые социальный опыт изменяет экспрессию генов в иммунных клетках. Каждая из этих моделей фокусируется на определённом аспекте, но вместе они создают мозаику механистических связей между уровнями, которая конкретизирует абстрактный плюралистический тезис. Продолжение этой работы механистической конкретизации — центральная задача современной науки о стрессе.
Альтернативные интегративные подходы — сетевая теория психопатологии и исследовательские критерии доменов — предлагают дополнительные инструменты для плюралистического исследования. Сетевой подход к психопатологии моделирует симптомы как узлы в причинной сети, где каждая связь может быть биологической, психологической или социальной по своему механизму. Исследовательские критерии доменов организуют изучение психопатологии вокруг трансдиагностических конструктов, измеряемых на всех уровнях от генов до поведения. Оба подхода имеют свои ограничения — сетевой подход рискует атеоретичностью, критерии доменов недостаточно учитывают социальный контекст — но они расширяют методологический арсенал плюралистического исследования. Синтез этих подходов с биопсихосоциальной рамкой и механистическими моделями остаётся задачей для будущего.
Эпистемологический плюрализм не означает отсутствия критериев для оценки теорий и методов; он означает признание множественности легитимных перспектив при сохранении требований научной строгости к каждой из них. Теория на любом уровне должна быть внутренне согласованной, эмпирически тестируемой, фальсифицируемой, обладать объяснительной и предсказательной силой. Данные на любом уровне должны быть надёжными, воспроизводимыми, полученными адекватными методами. Плюрализм не освобождает от этих требований; он добавляет к ним требование интеграции — объяснения того, как знание на одном уровне соотносится со знанием на других уровнях, какие связи, ограничения, взаимодействия существуют между уровнями. Это усложняет, а не упрощает задачу науки, но только такое усложнение адекватно сложности изучаемого феномена.
Практическое воплощение плюрализма в исследовательской практике требует междисциплинарности, которая выходит за рамки простого сотрудничества специалистов разных дисциплин и предполагает интеграцию перспектив на концептуальном уровне. Молекулярный биолог, психолог и социолог, изучающие один и тот же феномен стресса, могут работать изолированно, публикуя в разных журналах и не читая работ друг друга; это мультидисциплинарность без интеграции. Междисциплинарность предполагает, что они разрабатывают общие концептуальные рамки, формулируют гипотезы о связях между уровнями, планируют исследования, включающие переменные всех уровней, интерпретируют результаты в интегративной перспективе. Это требует преодоления дисциплинарных барьеров — языковых, институциональных, карьерных — что остаётся вызовом для академической системы, организованной по дисциплинарному принципу.
Связь эпистемологического плюрализма с образовательными целями данного курса фундаментальна и определяет его структуру и педагогику. Курс организован как последовательное рассмотрение уровней анализа стресса — от молекулярного и клеточного через системный к психологическому и социальному — не для того, чтобы представить их как изолированные, а для того, чтобы систематически изложить знание на каждом уровне с постоянным указанием на связи с другими уровнями. Сквозные связи между модулями, отмеченные в структуре курса, материализуют интегративный принцип: биологические механизмы связываются с психологическими процессами, психологические процессы — с социальными контекстами, патология — с интервенциями. Студент, прошедший курс, должен не только знать содержание каждого уровня, но и уметь мыслить вертикально — связывать уровни в понимании конкретных феноменов, переходить между уровнями объяснения, интегрировать разнородную информацию.
Когнитивные требования плюралистического мышления высоки и представляют педагогический вызов, который настоящий урок призван адресовать, закладывая эпистемологический фундамент для последующего обучения. Удержание в уме одновременно нескольких уровней анализа, понимание их связей и различий, способность переключаться между перспективами и интегрировать их — всё это требует когнитивной гибкости, которая не формируется автоматически при последовательном изучении отдельных дисциплин. Настоящий урок, посвящённый философским и методологическим основаниям, создаёт концептуальную карту, которая направляет последующее обучение и помогает позиционировать содержание каждого модуля в общей картине. Рефлексия над эпистемологическими вопросами — что мы можем знать, какими методами, какие ограничения, как интегрировать — формирует метакогнитивные навыки, необходимые для самостоятельного плюралистического мышления.
Завершая обсуждение эпистемологического плюрализма как необходимости для науки о стрессе, следует подчеркнуть, что эта позиция не является произвольным философским выбором, а вытекает из природы изучаемого феномена и из эмпирического опыта ограничений монистических подходов. Редукционизм генерирует ценное знание о механизмах, но систематически недооценивает эмерджентность и контекстуальность. Биомедицинская модель лечит болезни органов, но упускает целостность человека в контексте жизни. Психосоциальные подходы видят человека и контекст, но иногда минимизируют биологическую реальность. Трансляционные неудачи демонстрируют, что изолированное знание на одном уровне не транслируется в терапевтический успех. Все эти ограничения конвергируют к необходимости плюрализма — не как компромисса между крайностями, а как единственной позиции, адекватной сложности стресса как многоуровневого биопсихосоциального феномена. Этот урок закладывает фундамент для такого понимания, и последующие модули курса его развивают и конкретизируют.
Методологическая скромность и триангуляция
Эпистемологический плюрализм, обоснованный в предыдущем разделе, влечёт за собой определённую интеллектуальную установку, которую можно обозначить как методологическую скромность — признание того, что ни один отдельный уровень анализа, ни один исследовательский метод, ни одна теоретическая перспектива не способны предоставить полную и окончательную картину такого сложного феномена, как стресс. Эта скромность не является выражением агностицизма или релятивизма; она представляет собой рефлексивное осознание ограничений каждого исследовательского подхода, вытекающее из понимания многоуровневой природы изучаемого явления и из накопленного опыта неудач монистических стратегий. Методологическая скромность предполагает готовность признавать неполноту собственного знания, открытость к альтернативным объяснениям, внимание к данным, которые не укладываются в привычные рамки. Вместе с тем она не означает паралича или отказа от обобщений; она означает осторожность в формулировках, внимание к границам применимости выводов, постоянное стремление к расширению и уточнению понимания через интеграцию данных из множественных источников.
Концепция триангуляции предоставляет методологическую стратегию, позволяющую преодолевать ограничения отдельных методов и уровней анализа через их систематическое комбинирование. Термин заимствован из геодезии, где триангуляция означает определение положения точки по измерениям из нескольких известных позиций; в методологии исследований он обозначает стратегию использования множественных источников данных, методов, теорий или исследователей для изучения одного феномена. Логика триангуляции основана на том, что каждый метод и каждый уровень анализа имеет свои специфические ограничения и потенциальные источники искажений, но эти ограничения не совпадают между методами и уровнями. Если разные методы с разными паттернами ограничений конвергируют к одному и тому же выводу, вероятность того, что все они ошибаются одинаковым образом, снижается, и уверенность в выводе возрастает. Триангуляция не гарантирует истинности, но она повышает робастность выводов и защищает от систематических искажений, специфичных для отдельных методов.
Применение триангуляции к науке о стрессе означает систематическое сочетание данных с молекулярного, клеточного, системного, психологического и социального уровней для выявления закономерностей, которые воспроизводятся через эти уровни. Гипотеза о роли воспаления в депрессии, например, триангулируется через данные разных уровней: молекулярные исследования показывают связь провоспалительных цитокинов с нейротрансмиттерными системами; клеточные исследования демонстрируют активацию микроглии при хроническом стрессе; системные исследования выявляют повышенные воспалительные маркеры в крови пациентов с депрессией; психологические исследования показывают связь воспаления с ангедонией и когнитивными симптомами; эпидемиологические исследования обнаруживают коморбидность депрессии с воспалительными заболеваниями. Конвергенция данных со всех этих уровней создаёт значительно более прочную основу для воспалительной гипотезы, чем любой отдельный уровень мог бы предоставить. Если бы молекулярные данные указывали на роль воспаления, но клинические исследования не обнаруживали связи, это было бы сигналом о необходимости пересмотра гипотезы или её ограничений.
Триангуляция методов внутри одного уровня анализа представляет собой первый слой стратегии, позволяющий компенсировать ограничения конкретных экспериментальных техник и измерительных инструментов. На биологическом уровне изучение роли определённого гена в стрессовом ответе может включать генетические ассоциативные исследования у людей, эксперименты с нокаутными и трансгенными животными, клеточные исследования с манипуляцией экспрессии гена, анализ экспрессии в посмертных образцах мозга. Каждый из этих методов имеет свои ограничения: генетические ассоциации корреляционны и не доказывают причинность; животные модели не полностью переносимы на людей; клеточные модели теряют системный контекст; посмертные исследования отражают конечную стадию и подвержены артефактам. Однако если все эти методы указывают на одну и ту же роль гена, это существенно повышает уверенность. На психологическом уровне аналогичная триангуляция может включать самоотчётные опросники, поведенческие эксперименты, экологическую моментальную оценку, психофизиологические измерения — каждый метод схватывает разные аспекты психологического феномена и имеет разные источники искажений.
Триангуляция уровней анализа, связывающая биологические, психологические и социальные данные, представляет более амбициозную задачу, но именно она реализует потенциал плюралистического подхода наиболее полно. Когда гипотеза о механизме стресса подтверждается данными на всех уровнях — молекулярным механизмом, нейробиологическим субстратом, психологическим процессом, социальной детерминацией — она приобретает статус хорошо обоснованной теории, а не изолированной находки на одном уровне. Модель аллостатической нагрузки, рассмотренная ранее, представляет пример такой многоуровневой триангуляции: она связывает социальные стрессоры, психологическую оценку, нейроэндокринные каскады, воспалительные процессы, метаболические изменения, сердечно-сосудистые последствия в единую цепь, каждое звено которой подтверждается данными соответствующего уровня. Эта многоуровневая конвергенция создаёт убедительность, недостижимую для исследований на одном уровне. Вместе с тем достижение такой конвергенции требует масштабных междисциплинарных усилий и встречается в науке о стрессе реже, чем хотелось бы.
Триангуляция популяций представляет ещё одно измерение стратегии, особенно важное в контексте обсуждавшейся ранее гетерогенности человеческих популяций. Закономерность, обнаруженная в одной популяции, может не воспроизводиться в другой в силу генетических, культурных, средовых различий; только воспроизведение в множественных популяциях создаёт уверенность в универсальности или определяет границы применимости. Связь между детской травмой и депрессией во взрослом возрасте, например, воспроизводится в исследованиях разных стран, культур, социально-экономических контекстов, что указывает на робастность этой связи. Однако некоторые закономерности могут быть культурно специфичны: проявления стресса, копинг-стратегии, социальные детерминанты варьируют между культурами, и закономерность, верная для одного контекста, может не переноситься на другой. Триангуляция популяций позволяет различить универсальные механизмы и контекст-зависимые проявления, что критично для разработки интервенций, применимых глобально или требующих локальной адаптации.
Интерпретация конвергенции данных из разных источников представляет эпистемологическую задачу, требующую понимания того, что именно конвергенция означает и какую уверенность она предоставляет. Когда данные из независимых источников указывают на один и тот же механизм, это повышает вероятность того, что механизм реален, а не является артефактом конкретного метода или выборки. Однако конвергенция не является абсолютным доказательством: возможны систематические смещения, общие для разных методов, или ложные конвергенции, возникающие из-за недостаточной независимости источников. Критическая оценка конвергенции должна включать анализ того, насколько действительно независимы источники, какие смещения для них общие, какие альтернативные объяснения возможны. Методологическая скромность означает признание того, что даже конвергентные данные могут быть ревизованы в свете новых открытий, и что уверенность никогда не достигает абсолютной определённости. Наука прогрессирует через последовательное приближение, а не через окончательное доказательство.
Расхождение данных между уровнями или методами представляет не менее важную информацию, чем конвергенция, поскольку оно указывает на пробелы в понимании, ошибки в предположениях или необходимость уточнения теории. Когда молекулярные данные предсказывают определённый эффект, а клинические исследования его не обнаруживают, это расхождение требует объяснения. Возможные объяснения множественны: молекулярный механизм может действовать, но компенсироваться другими механизмами на системном уровне; клинические исследования могут быть недостаточно мощными или использовать неадекватные меры; популяция клинических исследований может отличаться от контекста, в котором механизм релевантен; теория, связывающая молекулярный и клинический уровни, может быть неполной. Каждое из этих объяснений генерирует гипотезы для дальнейшего исследования. Расхождение данных — не повод для отчаяния или игнорирования неудобных результатов; это сигнал о границах текущего понимания и направление для его расширения.
Трансляционный разрыв, детально обсуждавшийся ранее в данном уроке, представляет собой систематический паттерн расхождения между доклиническими и клиническими данными, который стимулировал рефлексию над методологическими предположениями доклинической науки. Когда препарат, эффективный на животной модели, не работает у людей, это расхождение указывает на ограничения модели, различия между видами, гетерогенность человеческих популяций, потерю контекста при трансляции. Накопление таких расхождений привело к осознанию необходимости более тщательной валидации моделей, использования транслируемых биомаркеров, стратификации пациентов, обратной трансляции. Все эти стратегии преодоления разрыва воплощают методологическую скромность — признание того, что первоначальные успехи в модельных системах не гарантируют клинического успеха и что необходима триангуляция через всё более приближённые к клинике источники данных.
Ограничения животных моделей, обсуждавшиеся подробно, иллюстрируют необходимость методологической скромности применительно к одному из центральных инструментов доклинической науки. Животные модели обеспечивают экспериментальный контроль, возможность манипуляции, доступ к биологическим образцам, которые невозможны у людей, и потому остаются незаменимыми для определённых исследовательских целей. Однако межвидовые различия в организации нервной системы, невозможность моделирования сложных когнитивных и эмоциональных феноменов человека, проблемы антропоморфизма создают систематические ограничения переносимости результатов. Методологически скромное отношение к животным данным означает их рассмотрение как гипотез для проверки на людях, а не как доказательств клинической эффективности. Триангуляция с человеческими данными — из клеточных моделей с человеческими клетками, исследований у здоровых добровольцев, клинических наблюдений — необходима для перехода от животных находок к терапевтическим приложениям.
Ограничения нейровизуализационных методов представляют ещё один пример области, требующей методологической скромности в интерпретации результатов. Функциональная магнитно-резонансная томография произвела революцию в изучении мозга при стрессе, позволив неинвазивно визуализировать активность мозговых областей и сетей. Однако интерпретация результатов нейровизуализации сопряжена с множеством ограничений: сигнал отражает не нейронную активность непосредственно, а связанные с ней метаболические изменения с задержкой в секунды; пространственное разрешение ограничено миллиметрами при нейронах микрометрового масштаба; статистический анализ включает множественные сравнения с риском ложноположительных результатов; воспроизводимость индивидуальных паттернов ограничена. Методологически скромное отношение к нейровизуализационным данным означает осторожность в локализационных выводах, внимание к размерам эффектов, требование воспроизведения, интеграцию с данными других методов — электрофизиологии, поражений, нейростимуляции.
Ограничения эпидемиологических методов, центральных для изучения социальных детерминант стресса и здоровья, также требуют методологической скромности. Эпидемиология оперирует большими выборками и позволяет выявлять связи на популяционном уровне, недоступные лабораторным исследованиям с их малыми выборками. Однако большинство эпидемиологических исследований наблюдательные и устанавливают корреляции, а не причинность. Конфаундеры — переменные, связанные и с предполагаемой причиной, и со следствием — могут создавать ложные ассоциации или маскировать реальные. Обратная причинность — когда следствие предшествует причине — особенно проблематична в кросс-секционных исследованиях. Смещение отбора и измерения искажают оценки связей. Методологически скромное отношение к эпидемиологическим данным означает осторожность в каузальных интерпретациях, внимание к потенциальным конфаундерам, использование методов каузальной инференции — менделевской рандомизации, естественных экспериментов, лонгитюдного моделирования — для укрепления каузальных выводов.
Ограничения самоотчётных методов в психологии стресса заслуживают отдельного рассмотрения в контексте методологической скромности. Шкалы воспринимаемого стресса, опросники симптомов, оценки жизненных событий составляют основу психологического измерения стресса и его последствий. Их преимущества очевидны: они доступны, дёшевы, схватывают субъективную перспективу, которая недоступна объективным измерениям. Однако самоотчёт подвержен множеству искажений: социальная желательность, ретроспективные искажения памяти, различия в интерпретации вопросов, текущее настроение, влияющее на оценку прошлого. Корреляции между воспринимаемым стрессом и биологическими маркерами часто умеренные или слабые, что ставит вопрос о том, что именно измеряют субъективные шкалы и как они соотносятся с объективной физиологией. Триангуляция самоотчёта с поведенческими измерениями, экологической моментальной оценкой, физиологическими маркерами необходима для более полной картины субъективного и объективного стресса.
Ограничения омиксных подходов, обсуждавшихся в контексте мультиомиксной интеграции, также требуют критического осмысления при всём их революционном потенциале. Геномика, транскриптомика, протеомика, метаболомика генерируют данные беспрецедентного масштаба, но этот масштаб создаёт специфические проблемы. Множественные сравнения при тестировании тысяч переменных порождают высокий риск ложноположительных результатов даже при строгих коррекциях. Переобучение моделей машинного обучения на обучающих данных не гарантирует генерализации на новые данные. Биологическая интерпретация статистически идентифицированных паттернов часто затруднена. Воспроизводимость омиксных находок между исследованиями ограничена из-за технических различий в платформах, протоколах, анализе. Методологически скромное отношение к омиксным данным означает требование внешней валидации, осторожность в биологических интерпретациях, интеграцию с функциональными экспериментами, подтверждающими механистическую значимость идентифицированных паттернов.
Ограничения системно-биологических подходов, рассмотренные в соответствующем разделе, напоминают, что новые парадигмы не свободны от собственных ограничений и не заменяют методологическую скромность технологическим оптимизмом. Сетевой анализ может идентифицировать структурные паттерны в данных, но знание структуры сети не эквивалентно пониманию её динамики и функции. Вычислительная сложность ограничивает масштаб анализируемых сетей. Неполнота данных об интерактомах означает, что анализируемые сети представляют частичную картину реальных взаимодействий. Интерпретируемость результатов алгоритмов машинного обучения ограничена, и статистический паттерн не всегда транслируется в биологическое понимание. Признание этих ограничений не умаляет ценности системных подходов, но помещает их в контекст инструментов, дополняющих, а не заменяющих другие методы и уровни анализа.
Культивирование методологической скромности в научном сообществе представляет задачу, выходящую за рамки индивидуальной добродетели и требующую институциональных изменений в практике науки. Система публикаций с её смещением в сторону положительных и статистически значимых результатов создаёт стимулы для преувеличения уверенности и минимизации ограничений. Конкуренция за гранты и позиции поощряет завышенные обещания и революционную риторику. Дисциплинарная специализация ограничивает знакомство с методами и ограничениями других дисциплин, способствуя методологическому парохиализму. Преодоление этих институциональных барьеров требует реформ в политике журналов, критериях финансирования, структуре образования, культуре научного сообщества. Регистрация исследований до их проведения, требование публикации негативных результатов, открытый доступ к данным для независимой проверки, междисциплинарная подготовка — все эти элементы реформы направлены на создание среды, поощряющей скромность и триангуляцию.
Роль критики и скептицизма в научном процессе приобретает особое значение в контексте методологической скромности. Критическая оценка методов, предположений, интерпретаций — не негативная деятельность, а необходимый компонент научного прогресса, выявляющий ограничения и направляющий к их преодолению. Воспроизведение чужих результатов, попытки фальсификации гипотез, поиск альтернативных объяснений — всё это выражения научного скептицизма, который уравновешивает энтузиазм открытий. В науке о стрессе критика биомедицинской модели привела к биопсихосоциальному подходу; критика животных моделей стимулировала развитие человеческих клеточных систем и органоидов; критика категориальной диагностики породила трансдиагностические и сетевые подходы. Каждый этап критики открывал новые направления развития. Методологическая скромность включает готовность принимать критику собственных подходов и использовать её для их совершенствования.
Связь методологической скромности и триангуляции с образовательными целями курса проявляется в формировании критического мышления как необходимой компетенции специалиста по стрессу. Курс не только передаёт знания о механизмах стресса на разных уровнях, но и формирует способность критически оценивать эти знания: понимать, какими методами они получены, какие ограничения у этих методов, какова степень уверенности в выводах, какие альтернативные интерпретации возможны. Модуль о научной грамотности закладывает основы критического мышления через обсуждение иерархии доказательств, различия корреляции и причинности, критериев оценки исследований. Последующие модули применяют эти критерии к конкретным областям, постоянно указывая на степень доказанности утверждений через систему маркировки. Студент, прошедший курс, должен не только знать, что известно о стрессе, но и понимать, как это знание получено и какова его надёжность.
Практические импликации методологической скромности для клинической практики состоят в осторожности при переносе исследовательских результатов на индивидуальных пациентов и в готовности пересматривать подходы в свете накапливающихся данных. Клиницист, информированный методологически скромным образом, понимает, что результаты клинических испытаний отражают средние эффекты в выборках и не гарантируют эффекта у конкретного пациента. Он использует клинические суждения, индивидуализирующие стандартные протоколы, мониторирует ответ на лечение и готов модифицировать стратегию. Он осведомлён о пределах текущего знания и честен с пациентами относительно неопределённости. Он открыт к новым данным и способен интегрировать их в практику. Эти качества не противоположны экспертизе; они представляют её зрелую форму, признающую, что экспертиза включает знание границ собственного знания.
Завершая обсуждение методологической скромности и триангуляции, следует подчеркнуть их неразрывную связь с эпистемологическим плюрализмом, обоснованным ранее. Плюрализм утверждает необходимость множественных перспектив; методологическая скромность признаёт ограничения каждой отдельной перспективы; триангуляция предоставляет стратегию для их интеграции в более надёжное знание. Вместе они образуют целостную эпистемологическую позицию для науки о стрессе: признание многоуровневой природы феномена, осознание ограничений каждого метода и уровня, систематическое комбинирование множественных источников для достижения конвергенции, интерпретация расхождений как указаний на пробелы, постоянное стремление к расширению и уточнению понимания. Эта позиция составляет эпистемологический фундамент курса и определяет его подход к представлению знания о стрессе — не как набора окончательных истин, а как динамично развивающегося понимания, обоснованного множественными источниками и открытого к ревизии.
Практические импликации для курса
Рассмотренные в данном уроке философские и методологические основания многоуровневого исследования стресса имеют прямые практические следствия для организации курса и формирования компетенций студентов, составляющих его образовательные цели. Урок занимает особое положение в архитектуре курса: он не представляет конкретных биологических механизмов или клинических феноменов, а закладывает эпистемологический фундамент, определяющий, как следует понимать и интерпретировать содержание всех последующих модулей. Без этого фундамента студент рискует воспринять курс как последовательность несвязанных блоков информации — молекулярного, нейробиологического, психологического, социального — каждый из которых самодостаточен и независим от других. С этим фундаментом студент понимает, что каждый модуль представляет аналитический срез единого многоуровневого феномена, что материал каждого модуля связан с материалом других модулей конкретными механизмами, что полное понимание стресса требует интеграции всех срезов в когерентную картину. Эта интегративная установка трансформирует способ восприятия и обработки информации на протяжении всего курса.
Структура курса, организованная как последовательное рассмотрение уровней анализа от молекулярного до социального, отражает педагогическую необходимость систематического изложения материала, но не должна создавать иллюзию изолированности уровней. Первый модуль рассматривает организм как адаптивную систему, вводя концепции гомеостаза и аллостаза, клеточные механизмы и митохондриальную энергетику. Второй модуль фокусируется на нервной регуляции и мозге, включая архитектуру нервной системы, ключевые структуры, мозговые сети, нейропластичность. Третий модуль детально рассматривает нейроэндокринные оси, особенно гипоталамо-гипофизарно-надпочечниковую ось и симпатоадреналовую систему. Четвёртый модуль посвящён иммунитету, воспалению и метаболизму, включая психонейроиммунологию, микробиом, метаболическую психиатрию. Пятый модуль переходит к психологии стресса — теориям, когнитивным процессам, эмоциям, копингу. Последующие модули охватывают измерение, патологию, интервенции, контексты, передовые исследования. Эта последовательность создаёт логику изложения, но каждый модуль содержит явные указания на связи с другими модулями, материализуя принцип интеграции.
Сквозные связи между модулями, систематически обозначенные в структуре курса, представляют собой конкретизацию многоуровневого подхода и заслуживают особого внимания студентов. Каждая тема в каждом модуле сопровождается указаниями на связанные темы в других модулях: молекулярные механизмы связываются с системными проявлениями, биологические субстраты — с психологическими процессами, патофизиология — с терапевтическими мишенями. Например, обсуждение глюкокортикоидного рецептора в третьем модуле связывается с нейропластичностью во втором модуле, с воспалением в четвёртом, с когнитивными эффектами стресса в пятом, с интервенциями в девятом. Студент, следящий за этими связями, выстраивает не линейное, а сетевое понимание материала, где каждый элемент занимает определённое положение в общей картине и связан множественными нитями с другими элементами. Это сетевое понимание соответствует реальной организации биологических систем, которая также сетевая, а не линейная.
Формирование способности к вертикальному мышлению, связывающему разные уровни анализа, представляет одну из центральных образовательных целей курса. Вертикальное мышление в данном контексте означает способность прослеживать, как феномен на одном уровне связан с феноменами на других уровнях — как социальный стрессор транслируется в психологическое переживание, как переживание активирует нейробиологические каскады, как каскады вызывают молекулярные изменения, и наоборот — как молекулярные изменения проявляются в клеточных дисфункциях, как дисфункции нарушают системную регуляцию, как нарушение регуляции создаёт психологические симптомы, как симптомы влияют на социальное функционирование. Студент с развитым вертикальным мышлением видит эти межуровневые связи спонтанно при рассмотрении любого аспекта стресса; он задаёт вопросы о том, каковы биологические субстраты психологического феномена и каковы психологические проявления биологического механизма. Это вертикальное мышление противоположно дисциплинарной изоляции, при которой специалист видит только свой уровень и игнорирует другие.
Формирование способности к горизонтальному мышлению, интегрирующему знание в пределах одного уровня, представляет вторую существенную образовательную цель. Горизонтальное мышление означает понимание того, как различные компоненты одного уровня взаимодействуют друг с другом, образуя целостную систему. На биологическом уровне это понимание того, как нервная, эндокринная и иммунная системы интегрированы в единую нейроэндокриноиммунную сеть, как различные нейромедиаторные системы модулируют друг друга, как метаболические и воспалительные процессы переплетаются. На психологическом уровне это понимание того, как когнитивные оценки, эмоции, поведенческие стратегии взаимообусловлены и не функционируют изолированно. На социальном уровне это понимание того, как семейные, профессиональные, культурные, экономические контексты взаимодействуют в определении стрессовой нагрузки. Горизонтальное мышление предотвращает фрагментацию понимания внутри уровня и создаёт основу для его вертикальной интеграции с другими уровнями.
Одновременное удержание деталей и целого представляет когнитивную задачу особой сложности, которую курс стремится развить через определённую педагогическую стратегию. Понимание стресса требует детального знания конкретных механизмов — специфических сигнальных путей, определённых нейронных цепей, конкретных психологических процессов, определённых социальных факторов. Без этого детального знания понимание остаётся поверхностным и не позволяет ни объяснять конкретные феномены, ни разрабатывать точные интервенции. Вместе с тем детальное знание само по себе недостаточно; необходимо понимание того, как детали складываются в целое, какова общая картина, в которую вписываются конкретные механизмы. Удержание одновременно деталей и целого требует способности к переключению масштабов: от микроуровня конкретного молекулярного взаимодействия до макроуровня системной динамики, от индивидуального пациента до популяционных закономерностей. Курс развивает эту способность через постоянное движение между уровнями детализации, через резюмирующие разделы, связывающие детали с общей картиной, через интегрирующие обсуждения, подобные настоящему.
Понимание специфичности механизмов в сочетании с контекстуальностью их действия представляет ещё одно измерение сложности, которому курс уделяет особое внимание. Биологические механизмы стресса имеют определённую специфичность: конкретные гормоны связываются с конкретными рецепторами, запускают конкретные сигнальные каскады, вызывают конкретные эффекты. Эта специфичность создаёт основу для понимания и вмешательства: зная механизм, можно разработать целенаправленную интервенцию. Однако действие этих специфичных механизмов контекстуально: эффект кортизола зависит от состояния рецепторов, которое зависит от предшествующей истории стресса; эффект психотерапии зависит от терапевтического альянса, который зависит от культурного контекста. Одна и та же молекула или одна и та же интервенция даёт различные эффекты в различных контекстах. Понимание этого сочетания специфичности и контекстуальности предохраняет как от чрезмерного механистического упрощения, так и от релятивистского отрицания механизмов.
Когнитивные требования многоуровневого мышления высоки и не формируются автоматически при последовательном изучении отдельных дисциплин. Традиционное дисциплинарное образование формирует глубокую экспертизу в одной области, но часто не развивает способность к интеграции с другими областями. Биолог глубоко понимает молекулярные механизмы, но может не видеть их психологических и социальных контекстов. Психолог детально знает когнитивные процессы, но может игнорировать их биологические субстраты. Междисциплинарное мышление, которое курс стремится сформировать, требует не только знаний из разных областей, но и способности их интегрировать, видеть связи, переключаться между перспективами, синтезировать разнородную информацию. Эти метакогнитивные навыки развиваются через практику: через анализ конкретных случаев с многоуровневой перспективы, через обсуждение связей между модулями, через рефлексию над собственными концептуальными рамками и их ограничениями.
Система маркировки доказательности, введённая в курсе, представляет практический инструмент, реализующий принцип методологической скромности в представлении материала. Каждое утверждение о механизмах, эффектах, интервенциях сопровождается указанием на степень его доказанности: консенсус, подтверждённый метаанализами; вероятная закономерность, поддержанная качественными рандомизированными исследованиями; спорное положение с противоречивыми данными; предварительная гипотеза на основе пилотных исследований; псевдонаучное утверждение, не подтверждённое доказательствами. Эта маркировка делает эксплицитной степень уверенности в каждом утверждении и предохраняет от ошибочного восприятия всего материала как одинаково обоснованного. Студент учится различать уровни доказанности, понимать, что знание имеет структуру с ядром хорошо обоснованных положений и периферией менее определённых гипотез. Это понимание критично для практического применения знаний.
Критическое мышление как образовательная цель пронизывает весь курс и получает своё эпистемологическое обоснование в настоящем уроке. Критическое мышление означает способность анализировать аргументы, оценивать доказательства, выявлять предположения, идентифицировать ограничения, рассматривать альтернативы. В контексте науки о стрессе это означает способность критически оценивать исследования — их дизайн, методы, выборки, статистику, интерпретации; понимать, что каждое исследование имеет ограничения и что выводы всегда условны; различать то, что данные показывают, и то, как их интерпретируют. Нулевой модуль о научной грамотности закладывает основы критического мышления, обучая критериям оценки исследований, различению корреляции и причинности, распознаванию псевдонауки. Последующие модули применяют эти критерии к конкретному содержанию, постоянно моделируя критический подход к оценке утверждений.
Иммунитет к псевдонауке и мифам о стрессе представляет практически важный результат эпистемологической подготовки. Область стресса особенно подвержена псевдонаучным утверждениям и мифам: «надпочечниковая усталость» как популярный, но научно не обоснованный диагноз; преувеличенные обещания отдельных добавок или практик; упрощённые объяснения вроде «стресс вызывает все болезни»; маркетинговая эксплуатация научной терминологии. Студент, прошедший курс, должен уметь распознавать эти псевдонаучные утверждения, понимать, почему они неверны или преувеличены, объяснять это пациентам и коллегам. Этот иммунитет основывается на понимании того, как устроено научное знание, какие критерии доказательности применяются, какая разница между проверенными механизмами и спекулятивными гипотезами. Обсуждение мифов и псевдонауки в нулевом модуле обязательно именно потому, что формирует этот критический иммунитет с самого начала обучения.
Формирование профессиональной идентичности специалиста по стрессу как интегративного мыслителя представляет долгосрочную цель курса, выходящую за рамки передачи конкретных знаний. Специалист по стрессу — не просто биолог, психолог или клиницист, применяющий свою дисциплину к теме стресса; это профессионал с интегративным видением, способный работать на пересечении дисциплин, координировать мультидисциплинарные команды, переводить между профессиональными языками, синтезировать разнородную информацию для решения конкретных проблем. Такая профессиональная идентичность формируется не только через усвоение знаний, но и через присвоение определённого способа мышления, который данный урок артикулирует и обосновывает. Студент, идентифицирующий себя как интегративного специалиста, естественно ищет связи между уровнями, признаёт ограничения своей экспертизы, открыт к сотрудничеству с коллегами из других областей.
Применение многоуровневого понимания к клинической практике составляет важнейшую практическую цель для студентов, ориентированных на работу с пациентами. Клиницист с многоуровневым мышлением оценивает пациента не только с точки зрения симптомов и диагноза, но и в контексте его биологической конституции, психологических особенностей, социальной ситуации. Он формулирует случай в биопсихосоциальных терминах, идентифицируя вклад факторов каждого уровня и их взаимодействия. Он планирует интервенции, учитывающие множественные уровни: биологические, когда это показано; психологические, адресующие когнитивные и эмоциональные процессы; социальные, мобилизующие поддержку и изменяющие контекст. Он мониторирует ответ на лечение на нескольких уровнях и готов модифицировать стратегию в свете получаемой информации. Это многоуровневое клиническое мышление развивается на протяжении всего курса, особенно в модулях о патологии, интервенциях и контекстах, но его концептуальная основа закладывается в настоящем уроке.
Применение многоуровневого понимания к исследовательской практике определяет, как студенты, ориентированные на научную карьеру, будут планировать и проводить собственные исследования. Исследователь с многоуровневым мышлением формулирует вопросы, учитывающие связи между уровнями, а не изолированные закономерности на одном уровне. Он планирует исследования, включающие измерения на нескольких уровнях, позволяющие тестировать гипотезы о межуровневых связях. Он интерпретирует результаты в контексте того, что известно на других уровнях, ища конвергенцию или объясняя расхождения. Он осознаёт ограничения своих методов и ищет триангуляцию с методами из других традиций. Он сотрудничает с коллегами из других дисциплин, чья экспертиза дополняет его собственную. Всё это отражает применение эпистемологических принципов, артикулированных в данном уроке, к конкретной исследовательской практике.
Применение многоуровневого понимания к разработке интервенций и политик расширяет практические импликации за пределы индивидуальной работы с пациентами или исследовательскими проектами. Разработчик интервенций с многоуровневым мышлением рассматривает воздействия на разных уровнях — фармакологические, психотерапевтические, поведенческие, социальные, средовые — и ищет их оптимальную комбинацию для конкретной проблемы и популяции. Он понимает, что интервенция на одном уровне имеет эффекты на других уровнях через межуровневые связи. Он проектирует оценку интервенций, включающую исходы на нескольких уровнях. Разработчик политик с многоуровневым мышлением понимает, как макроуровневые решения — экономическая политика, организация здравоохранения, городское планирование — транслируются через мезоуровень организаций и сообществ на микроуровень индивидуального здоровья. Это системное понимание политических решений следует из многоуровневой концептуализации стресса.
Непрерывное обучение и профессиональное развитие после завершения курса также определяются сформированным эпистемологическим фундаментом. Наука о стрессе динамично развивается: появляются новые методы, открываются новые механизмы, пересматриваются прежние представления. Специалист, сформированный курсом, должен быть способен интегрировать новую информацию в своё понимание, критически оценивать новые утверждения, обновлять практику в свете новых данных. Эпистемологический фундамент — понимание того, как устроено научное знание, какова его структура и динамика — создаёт основу для этого непрерывного обучения. Он позволяет не просто запоминать факты, которые могут устареть, а понимать принципы, которые остаются применимы к оценке новых фактов. Метакогнитивные навыки критического мышления, многоуровневой интеграции, методологической скромности переносимы на новый материал и служат инструментами самостоятельного профессионального развития.
Курс как целое представляет собой воплощение многоуровневого подхода к пониманию стресса, и завершающая рефлексия над его структурой помогает студентам увидеть эту архитектуру эксплицитно. От нулевого модуля, закладывающего язык и критическое мышление, через биологические модули, раскрывающие механизмы от молекулы до системы, через психологический модуль, освещающий субъективное измерение, через модуль измерения, связывающий концептуальное с эмпирическим, через модули о патологии и интервенциях, применяющие понимание к клинической реальности, к модулю о контекстах, расширяющему перспективу на популяции, и модулю о передовых исследованиях, открывающему горизонт развития — курс выстраивает многослойное понимание стресса, где каждый слой связан с другими. Эта архитектура не произвольна; она отражает структуру самого феномена и воплощает эпистемологические принципы, обоснованные в данном уроке.
Настоящий урок, завершаясь, переводит студента в состояние готовности к полноценному усвоению последующего материала с интегративной установкой, которая трансформирует способ его восприятия. Без этой подготовки студент рисковал бы воспринять биологические модули как чистую биологию, психологический модуль как чистую психологию, и так далее — фрагментированно и изолированно. С этой подготовкой студент воспринимает каждый модуль как срез единого феномена, ищет связи с другими срезами, формирует интегрированное понимание. Эпистемологический фундамент — не абстрактная философия, не имеющая практического значения; это концептуальная карта, определяющая, как располагается и связывается весь последующий материал. Студент, усвоивший этот урок, готов к погружению в конкретные механизмы стресса с пониманием их места в общей картине и с навыками, необходимыми для самостоятельной интеграции знаний.
Завершая интегрирующее обсуждение и урок в целом, следует подчеркнуть, что эпистемологический фундамент, заложенный здесь, является не завершённой конструкцией, а отправной точкой для развития, которое продолжается на протяжении всего курса и профессиональной жизни. Философские и методологические принципы — эпистемологический плюрализм, методологическая скромность, триангуляция, вертикальное и горизонтальное мышление — не усваиваются раз и навсегда при первом чтении; они интернализируются через повторное применение к конкретному материалу, через рефлексию над собственным мышлением, через столкновение с реальными проблемами понимания и интервенции. Курс предоставляет множество возможностей для такого применения и рефлексии; студент должен использовать их активно, превращая эпистемологические принципы из абстрактных формулировок в интернализованные интеллектуальные привычки. Это трудная когнитивная задача, но она необходима для формирования современного специалиста по стрессу, способного понимать, исследовать и модифицировать этот сложный многоуровневый феномен.
Практикум
Для базового слоя
PDF формы задания
Пятиуровневый разбор реакции на нагрузку
Разберите один эпизод: контекст, оценки, телесные реакции и связи уровней, чтобы выбрать точки влияния.
Диагностика дисфункций и ресурсов по уровням
Сравнение признаков нарушений и ресурсов на 5 уровнях с выбором приоритетов и интегрированным планом помощи.
Пирамида пяти уровней и связей причинности
Схема уровней от молекул до социума: направления влияния, ошибки редукции и пример разбора.
Проверка понимания причинности между уровнями
Вопросы на связи уровней: выбор ответов, сопоставление, кейсы и проектирование комплексной помощи.
Интерактивные формы - задания
Интерактивная схема-памятка пяти уровней анализа стресса: от молекул до социума. Изучите каждый уровень и увидите, как один стрессор «увольнение» проявляется на всех этажах пирамиды.
Распределите 25 карточек по пяти уровням анализа стресса: молекулярный, клеточный, системный, психологический и социальный. Проверьте понимание многоуровневого подхода к изучению стресса.
Пошаговый разбор реального стрессора через призму пяти уровней анализа — от социального до молекулярного. Определите релевантные проявления на каждом уровне и установите причинно-следственные связи.
Соедините 10 терминов стресс-реакции с соответствующими уровнями анализа (молекулярный, клеточный, системный, психологический, социальный), а затем с примерами из ситуации увольнения. Система проверит точность и объяснит ошибки.
Постройте причинную цепочку из 7 звеньев, связывающую социальный стрессор с молекулярными изменениями. Проверьте многоуровневое мышление: от увольнения через психологию и физиологию к клеткам.
Разложите свой недавний стресс по пяти уровням анализа: от социального контекста до молекулярных процессов. Рефлексивное упражнение поможет увидеть привычные «слепые зоны» и наметить конкретное действие.
Проверьте понимание многоуровневого подхода к стрессу. 10 вопросов: определите, к какому уровню относится феномен, и как уровни связаны между собой. Порог прохождения — 7 из 10 баллов.
Прочитайте описание случая и определите, к какому уровню анализа относится каждая фраза: молекулярному, клеточному, системному, психологическому или социальному. Научитесь видеть вертикальный разрез стрессовой реакции.
Для академического слоя
PDF формы - задания
Критический анализ статьи по охвату уровней
Оценка охвата и глубины уровней в публикации, изучение межуровневых связей, пробелов и улучшений дизайна.
Картирование восходящих и нисходящих связей
Построение многоуровневой модели феномена: процессы, механизмы, циклы, временная динамика и гипотезы.
Границы редукционизма в объяснениях
Разбор предпосылок и последствий редукции: потеря контекста, игнорирование причинности, альтернативные трактовки.
Дизайн многоуровневого исследования
Планирование вопроса, измерений и гипотез, временной структуры и аналитики с учётом ограничений.
Интерактивные формы - задания
Станьте рецензентом исследования о стрессе. Оцените 6 утверждений автора: когда редукционизм уместен, а когда требуется учёт эмерджентности, нисходящей причинности или множественной реализуемости.
Спроектируйте дизайн научного исследования стресса: выберите вопрос, постройте иерархию уровней анализа, операционализируйте переменные, задайте медиаторы и модераторы. В конце — экспорт готового плана.
Пройдите 6 этапов трансляции открытия — от клеточной культуры до внедрения. На каждом этапе определите ключевые риски и стратегии их снижения. В конце — карта слабых мест.
Постройте сеть взаимосвязей между компонентами стресс-реакции. Добавьте связи между узлами, найдите петли обратной связи и определите «рычажные точки» — места оптимального вмешательства в систему.
Классифицируйте 8 утверждений о биопсихосоциальной модели по категориям и обоснуйте каждый выбор. Практикум проверяет понимание сильных сторон модели и её ключевых ограничений.
Проанализируйте вымышленный абстракт исследования стресса. Определите, какие уровни анализа были измерены, какие пропущены, и какие «мосты» между уровнями отсутствуют. Оцените ограничения интерпретации.
Проанализируйте вымышленное исследование стресса на предмет методологических ограничений редукционистского подхода. Выберите ограничения, предложите улучшения дизайна и напишите мини-рецензию.
Соберите интегративную модель феномена, связав социальный, психологический и биологические уровни каузальными путями. Укажите механизмы влияния и критерии проверяемости модели.